

Functionally Diverse Inflammatory Responses in Peripheral and Liver Monocytes in Alcohol-Associated Hepatitis
- PMID: 33024916
- PMCID: PMC7527760
- DOI: 10.1002/hep4.1563
Functionally Diverse Inflammatory Responses in Peripheral and Liver Monocytes in Alcohol-Associated Hepatitis
Abstract
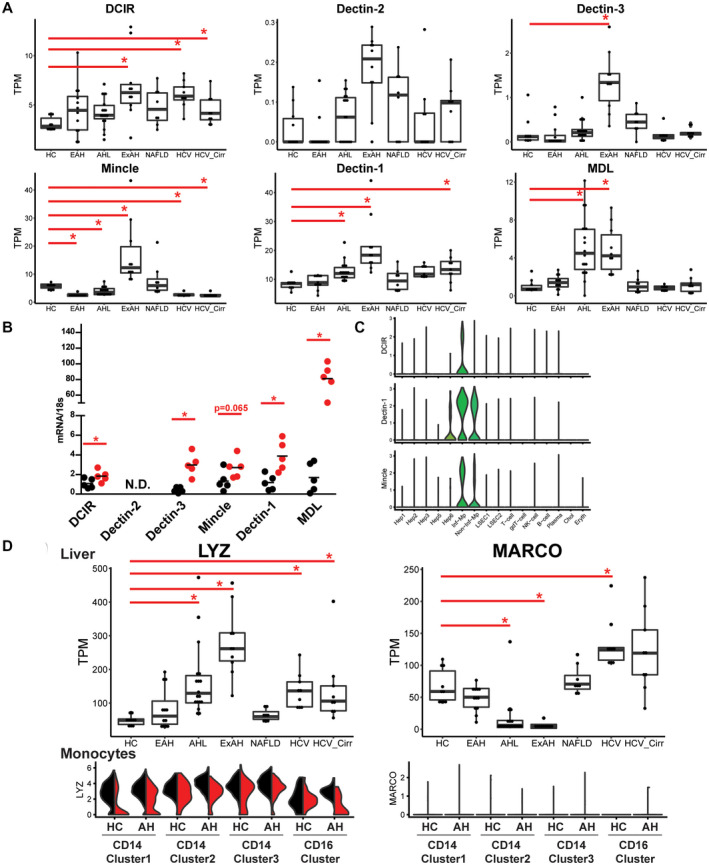
Alcohol-associated hepatitis (AH) is an acute inflammatory disease in which gut-microbial byproducts enter circulation and peripheral immune cells infiltrate the liver, leading to nonresolving inflammation and injury. Single-cell RNA sequencing of peripheral blood mononuclear cells isolated from patients with AH and healthy controls paired with lipopolysaccharide (LPS) challenge revealed how diverse monocyte responses are divided among individual cells and change in disease. After LPS challenge, one monocyte subtype expressed pro-inflammatory genes in both disease and healthy controls, while another monocyte subtype was anti-inflammatory in healthy controls but switched to pro-inflammatory in AH. Numerous immune genes are clustered within genomic cassettes, including chemokines and C-type lectin receptors (CTRs). CTRs sense byproducts of diverse microbial and host origin. Single-cell data revealed correlated expression of genes within cassettes, thus further diversifying different monocyte responses to individual cells. Monocyte up-regulation of CTRs in response to LPS caused hypersensitivity to diverse microbial and host-derived byproducts, indicating a secondary immune surveillance pathway up-regulated in a subset of cells by a closely associated genomic cassette. Finally, expression of CTR genes was higher in livers of patients with severe AH, but not other chronic liver diseases, implicating secondary immune surveillance in nonresolving inflammation in severe AH.
© 2020 The Authors. Hepatology Communications published by Wiley Periodicals LLC on behalf of American Association for the Study of Liver Diseases.
Figures






Similar articles
-
miRNAs Involved in M1/M2 Hyperpolarization Are Clustered and Coordinately Expressed in Alcoholic Hepatitis.Front Immunol. 2019 Jun 7;10:1295. doi: 10.3389/fimmu.2019.01295. eCollection 2019. Front Immunol. 2019. PMID: 31231396 Free PMC article.
-
Single-cell transcriptomics of peripheral blood mononuclear cells indicates impaired immune and inflammatory responses in alcohol-associated hepatitis.Hum Immunol. 2024 Jan;85(1):110735. doi: 10.1016/j.humimm.2023.110735. Epub 2023 Nov 30. Hum Immunol. 2024. PMID: 38040543
-
Genome Restructuring around Innate Immune Genes in Monocytes in Alcohol-associated Hepatitis.bioRxiv [Preprint]. 2024 Aug 9:2024.08.07.607014. doi: 10.1101/2024.08.07.607014. bioRxiv. 2024. PMID: 39211112 Free PMC article. Preprint.
-
Peripheral blood mononuclear cell mitochondrial dysfunction in acute alcohol-associated hepatitis.Clin Transl Med. 2023 May;13(5):e1276. doi: 10.1002/ctm2.1276. Clin Transl Med. 2023. PMID: 37228227 Free PMC article.
-
Decoding cell death signals in liver inflammation.J Hepatol. 2013 Sep;59(3):583-94. doi: 10.1016/j.jhep.2013.03.033. Epub 2013 Apr 6. J Hepatol. 2013. PMID: 23567086 Review.
Cited by
-
Role of MIF in coordinated expression of hepatic chemokines in patients with alcohol-associated hepatitis.JCI Insight. 2021 Jun 8;6(11):e141420. doi: 10.1172/jci.insight.141420. JCI Insight. 2021. PMID: 33945507 Free PMC article.
-
The pyruvate dehydrogenase kinase inhibitor dichloroacetate mitigates alcohol-induced hepatic inflammation and metabolic disturbances in mice.Hepatol Commun. 2024 Nov 29;8(12):e0547. doi: 10.1097/HC9.0000000000000547. eCollection 2024 Dec 1. Hepatol Commun. 2024. PMID: 39621302 Free PMC article.
-
Recent Advances in Understanding of Pathogenesis of Alcohol-Associated Liver Disease.Annu Rev Pathol. 2023 Jan 24;18:411-438. doi: 10.1146/annurev-pathmechdis-031521-030435. Epub 2022 Oct 21. Annu Rev Pathol. 2023. PMID: 36270295 Free PMC article. Review.
-
Crosstalk between Oxidative Stress and Inflammatory Liver Injury in the Pathogenesis of Alcoholic Liver Disease.Int J Mol Sci. 2022 Jan 11;23(2):774. doi: 10.3390/ijms23020774. Int J Mol Sci. 2022. PMID: 35054960 Free PMC article. Review.
-
Transcriptional and Epigenetic Regulation of Monocyte and Macrophage Dysfunction by Chronic Alcohol Consumption.Front Immunol. 2022 Jun 29;13:911951. doi: 10.3389/fimmu.2022.911951. eCollection 2022. Front Immunol. 2022. PMID: 35844518 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
