

Rosettes integrity protects Plasmodium vivax of being phagocytized
- PMID: 33028898
- PMCID: PMC7541459
- DOI: 10.1038/s41598-020-73713-w
Rosettes integrity protects Plasmodium vivax of being phagocytized
Abstract
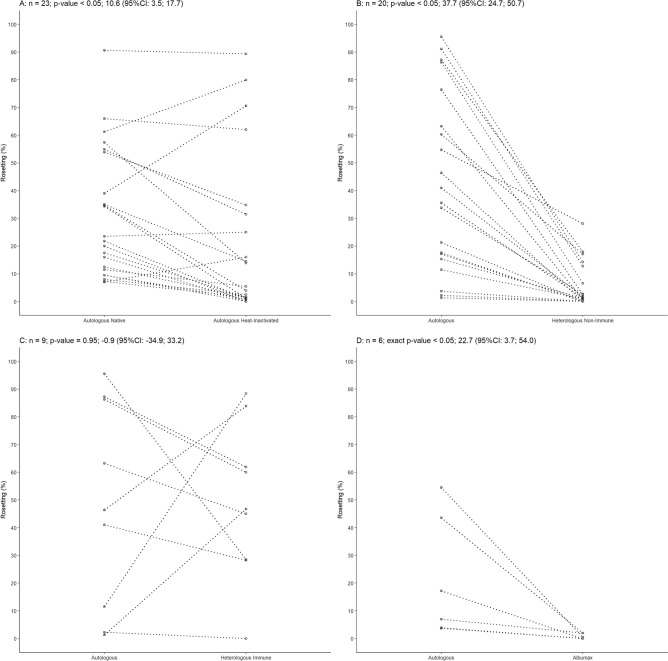
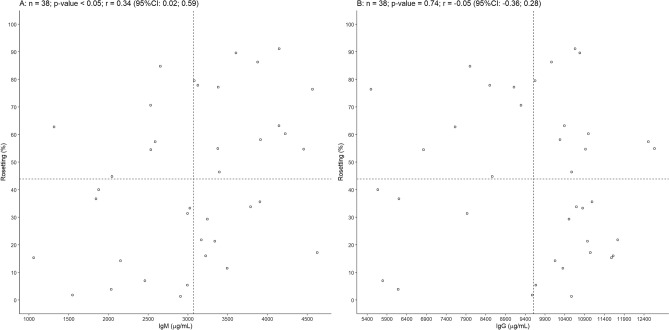
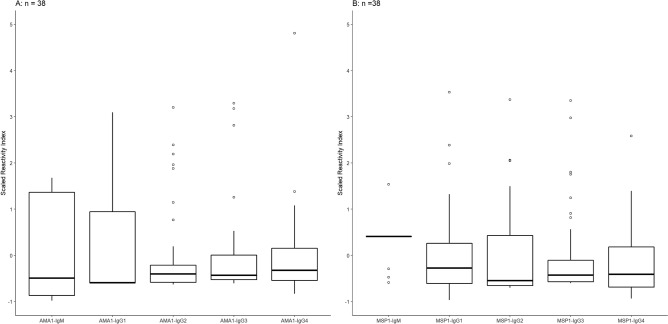
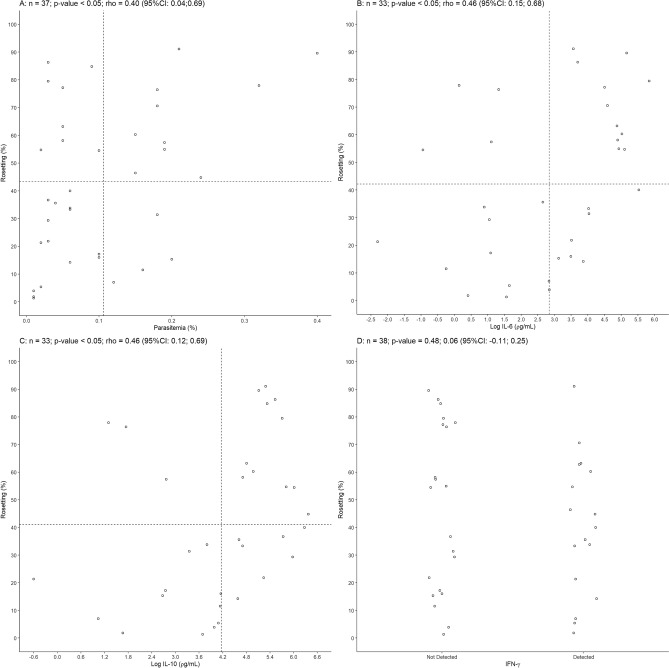
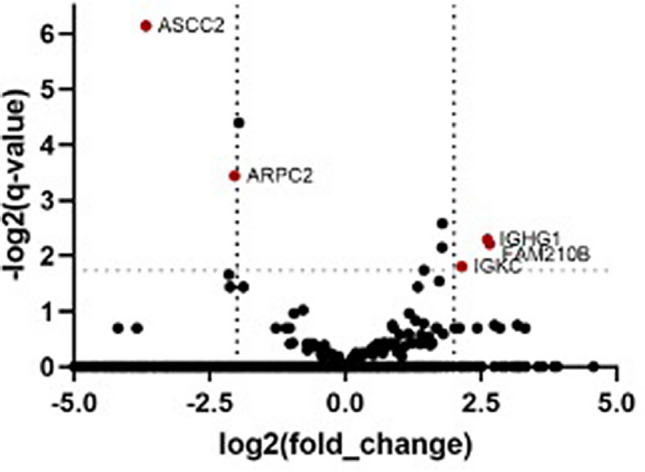
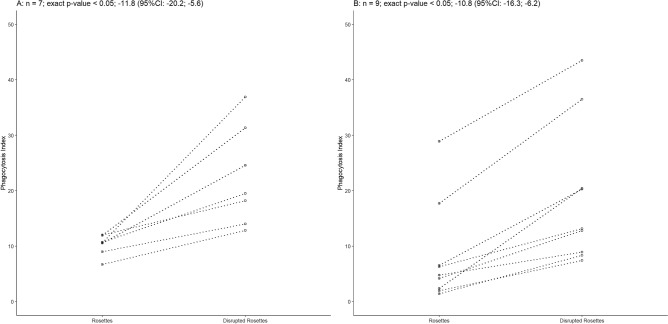
Plasmodium vivax is the most prevalent cause of malaria outside of Africa. P. vivax biology and pathogenesis are still poorly understood. The role of one highly occurring phenotype in particular where infected reticulocytes cytoadhere to noninfected normocytes, forming rosettes, remains unknown. Here, using a range of ex vivo approaches, we showed that P. vivax rosetting rates were enhanced by plasma of infected patients and that total immunoglobulin M levels correlated with rosetting frequency. Moreover, rosetting rates were also correlated with parasitemia, IL-6 and IL-10 levels in infected patients. Transcriptomic analysis of peripheral leukocytes from P. vivax-infected patients with low or moderated rosetting rates identified differentially expressed genes related to human host phagocytosis pathway. In addition, phagocytosis assay showed that rosetting parasites were less phagocyted. Collectively, these results showed that rosette formation plays a role in host immune response by hampering leukocyte phagocytosis. Thus, these findings suggest that rosetting could be an effective P. vivax immune evasion strategy.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
