

Knockout of PRDX6 induces mitochondrial dysfunction and cell cycle arrest at G2/M in HepG2 hepatocarcinoma cells
- PMID: 33035814
- PMCID: PMC7554216
- DOI: 10.1016/j.redox.2020.101737
Knockout of PRDX6 induces mitochondrial dysfunction and cell cycle arrest at G2/M in HepG2 hepatocarcinoma cells
Abstract
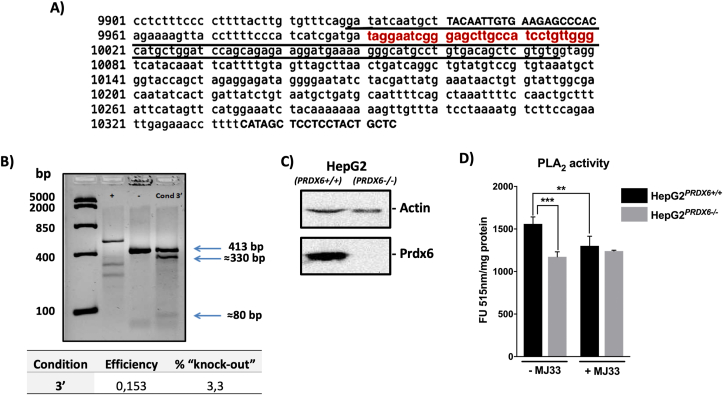
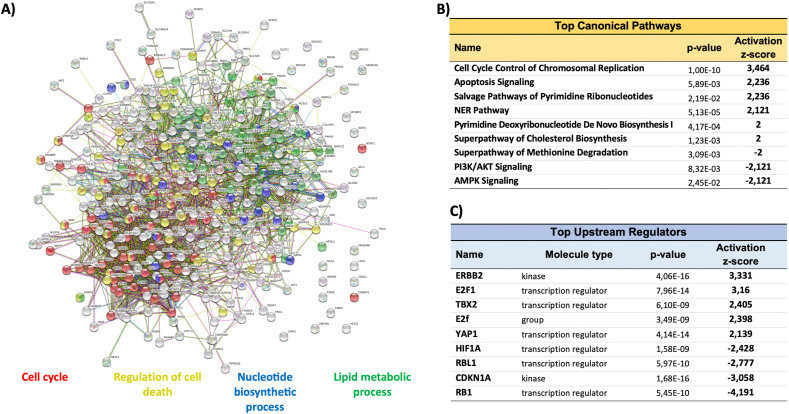
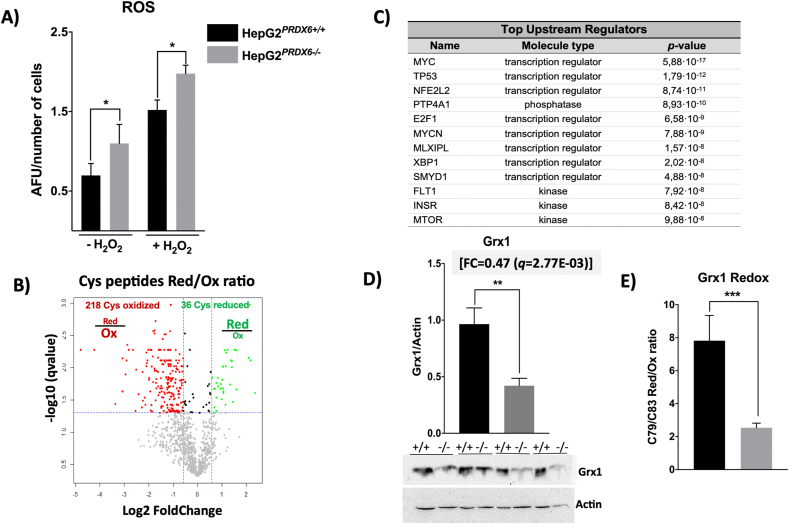
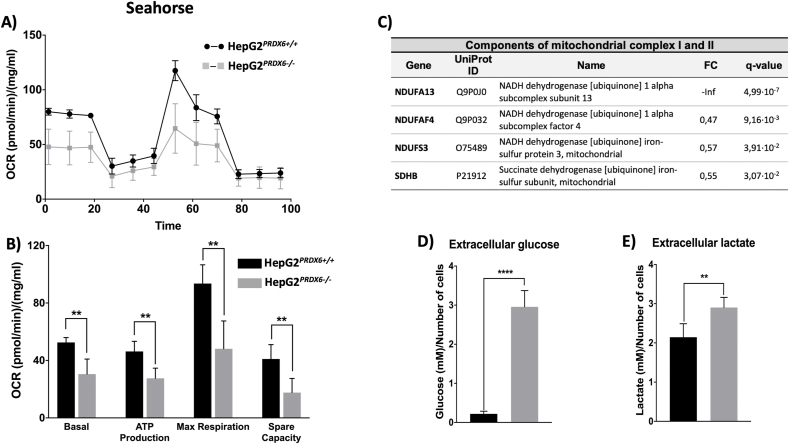
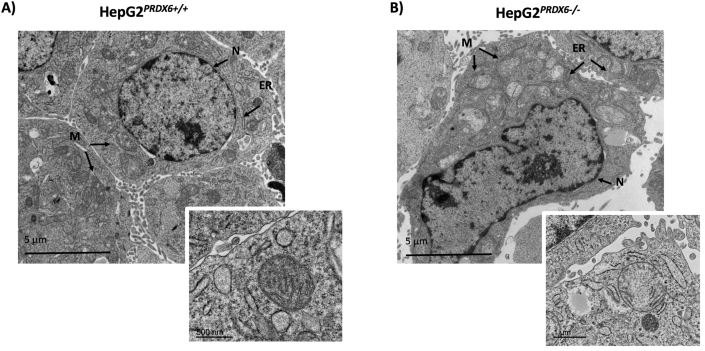
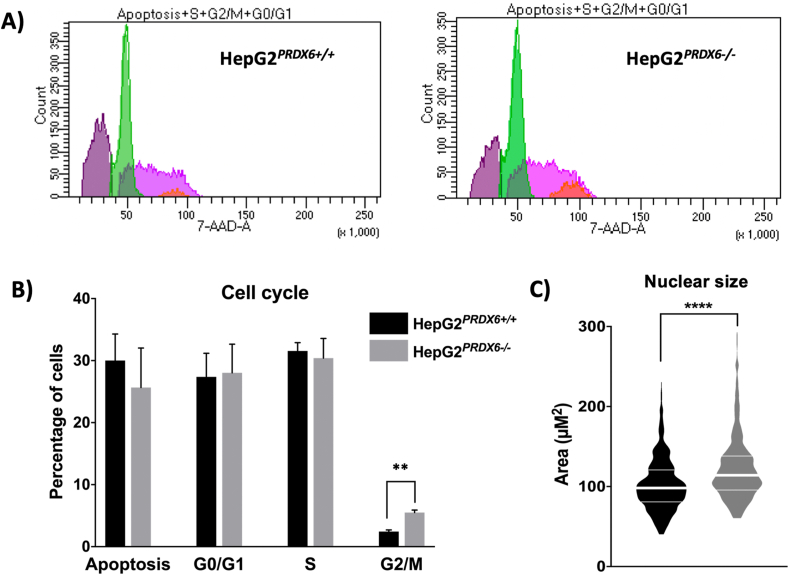
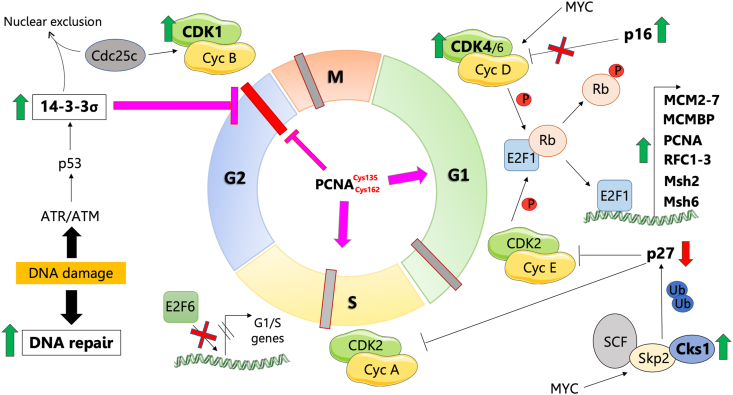
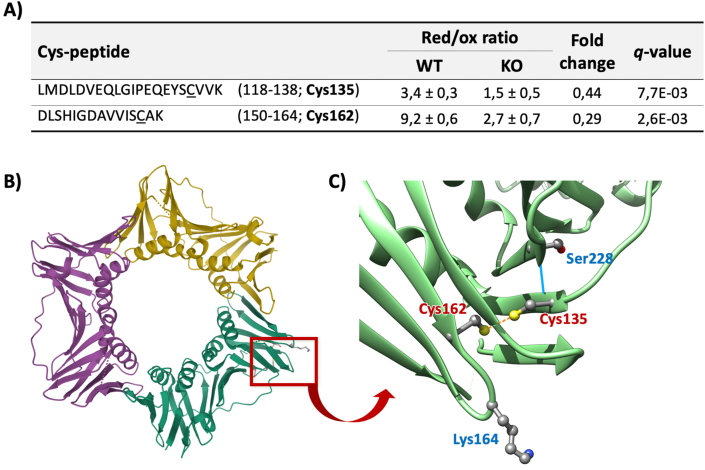
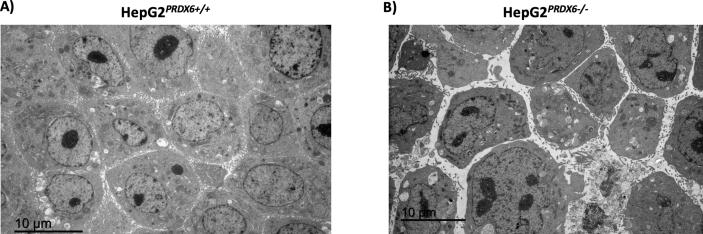
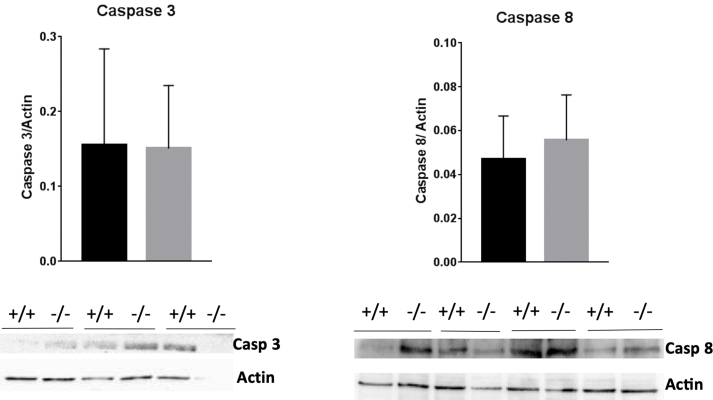
Peroxiredoxin 6 (PRDX6) has been associated with tumor progression and cancer metastasis. Its acting on phospholipid hydroperoxides and its phospholipase-A2 activity are unique among the peroxiredoxin family and add complexity to its action mechanisms. As a first step towards the study of PRDX6 involvement in cancer, we have constructed a human hepatocarcinoma HepG2PRDX6-/- cell line using the CRISPR/Cas9 technique and have characterized the cellular response to lack of PRDX6. Applying quantitative global and redox proteomics, flow cytometry, in vivo extracellular flow analysis, Western blot and electron microscopy, we have detected diminished respiratory capacity, downregulation of mitochondrial proteins and altered mitochondrial morphology. Autophagic vesicles were abundant while the unfolded protein response (UPR), HIF1A and NRF2 transcription factors were not activated, despite increased levels of p62/SQSTM1 and reactive oxygen species (ROS). Insulin receptor (INSR), 3-phosphoinositide-dependent protein kinase 1 (PDPK1), uptake of glucose and hexokinase-2 (HK2) decreased markedly while nucleotide biosynthesis, lipogenesis and synthesis of long chain polyunsaturated fatty acids (LC-PUFA) increased. 254 Cys-peptides belonging to 202 proteins underwent significant redox changes. PRDX6 knockout had an antiproliferative effect due to cell cycle arrest at G2/M transition, without signs of apoptosis. Loss of PLA2 may affect the levels of specific lipids altering lipid signaling pathways, while loss of peroxidase activity could induce redox changes at critical sensitive cysteine residues in key proteins. Oxidation of specific cysteines in Proliferating Cell Nuclear Antigen (PCNA) could interfere with entry into mitosis. The GSH/Glutaredoxin system was downregulated likely contributing to these redox changes. Altogether the data demonstrate that loss of PRDX6 slows down cell division and alters metabolism and mitochondrial function, so that cell survival depends on glycolysis to lactate for ATP production and on AMPK-independent autophagy to obtain building blocks for biosynthesis. PRDX6 is an important link in the chain of elements connecting redox homeostasis and proliferation.
Keywords: CRISPR-Cas9; Carbohydrate metabolism; Cell cycle; Glucose metabolism; Lipokines; Mitochondria; NRF2; PCNA; Peroxiredoxin 6; Proteomics; Redox proteome.
Copyright © 2020 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Peroxiredoxin 6 Down-Regulation Induces Metabolic Remodeling and Cell Cycle Arrest in HepG2 Cells.Antioxidants (Basel). 2019 Oct 23;8(11):505. doi: 10.3390/antiox8110505. Antioxidants (Basel). 2019. PMID: 31652719 Free PMC article.
-
Peroxiredoxin 6 is the primary antioxidant enzyme for the maintenance of viability and DNA integrity in human spermatozoa.Hum Reprod. 2018 Aug 1;33(8):1394-1407. doi: 10.1093/humrep/dey221. Hum Reprod. 2018. PMID: 29912414 Free PMC article.
-
Loss of peroxiredoxin 6 alters lipid composition and distribution resulting in increased sensitivity to ferroptosis.Biochem J. 2024 Dec 23;481(24):1997-2015. doi: 10.1042/BCJ20240445. Biochem J. 2024. PMID: 39601357 Free PMC article.
-
Peroxiredoxin 6, a 1-Cys peroxiredoxin, functions in antioxidant defense and lung phospholipid metabolism.Free Radic Biol Med. 2005 Jun 1;38(11):1422-32. doi: 10.1016/j.freeradbiomed.2005.02.011. Free Radic Biol Med. 2005. PMID: 15890616 Review.
-
Structural and Functional Diversity of the Peroxiredoxin 6 Enzyme Family.Antioxid Redox Signal. 2024 May;40(13-15):759-775. doi: 10.1089/ars.2023.0287. Epub 2023 Sep 12. Antioxid Redox Signal. 2024. PMID: 37463006 Review.
Cited by
-
Resveratrol and Its Analogs: Potent Agents to Reverse Epithelial-to-Mesenchymal Transition in Tumors.Front Oncol. 2021 Apr 16;11:644134. doi: 10.3389/fonc.2021.644134. eCollection 2021. Front Oncol. 2021. PMID: 33937049 Free PMC article. Review.
-
The Relationship of Redox With Hallmarks of Cancer: The Importance of Homeostasis and Context.Front Oncol. 2022 Apr 22;12:862743. doi: 10.3389/fonc.2022.862743. eCollection 2022. Front Oncol. 2022. PMID: 35530337 Free PMC article. Review.
-
Akt3 links mitochondrial function to the regulation of Aurora B and mitotic fidelity.PLoS One. 2025 Mar 6;20(3):e0315751. doi: 10.1371/journal.pone.0315751. eCollection 2025. PLoS One. 2025. PMID: 40048438 Free PMC article.
-
Hexokinase 2 promoted cell motility and proliferation by activating Akt1/p-Akt1 in human ovarian cancer cells.J Ovarian Res. 2022 Aug 11;15(1):92. doi: 10.1186/s13048-022-01027-8. J Ovarian Res. 2022. PMID: 35953860 Free PMC article.
-
α-Amanitin aggravates hepatic injury by activating oxidative stress and mitophagy via peroxiredoxin 6 inhibition.Immunol Res. 2025 Mar 19;73(1):64. doi: 10.1007/s12026-025-09619-4. Immunol Res. 2025. PMID: 40108092
References
-
- Pedrajas J.R., McDonagh B., Hernandez-Torres F., Miranda-Vizuete A., Gonzalez-Ojeda R., Martinez-Galisteo E., Padilla C.A., Barcena J.A. Glutathione is the resolving thiol for thioredoxin peroxidase activity of 1-cys peroxiredoxin without being consumed during the catalytic cycle. Antioxidants Redox Signal. 2016;24:115–128. doi: 10.1089/ars.2015.6366. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
