

ATP-Binding and Hydrolysis in Inflammasome Activation
- PMID: 33036374
- PMCID: PMC7583971
- DOI: 10.3390/molecules25194572
ATP-Binding and Hydrolysis in Inflammasome Activation
Abstract
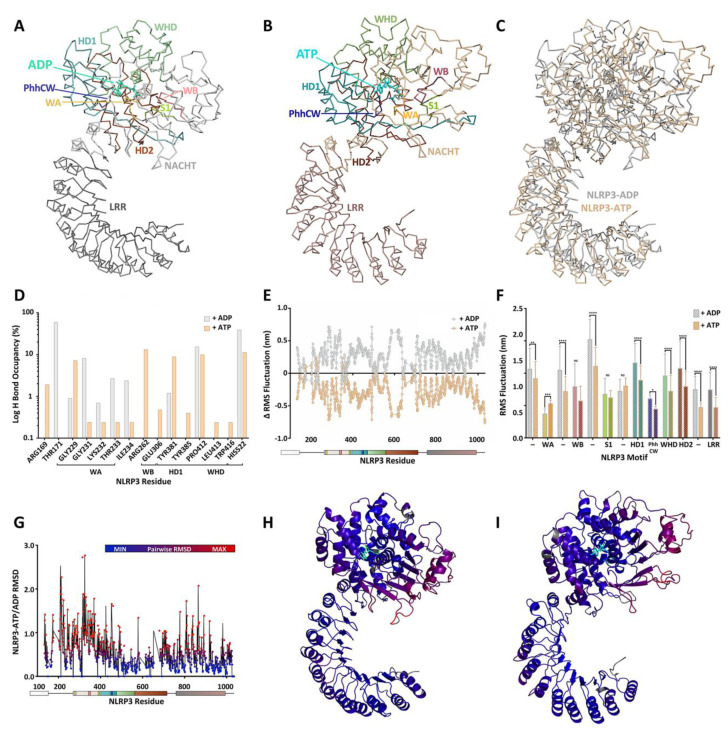
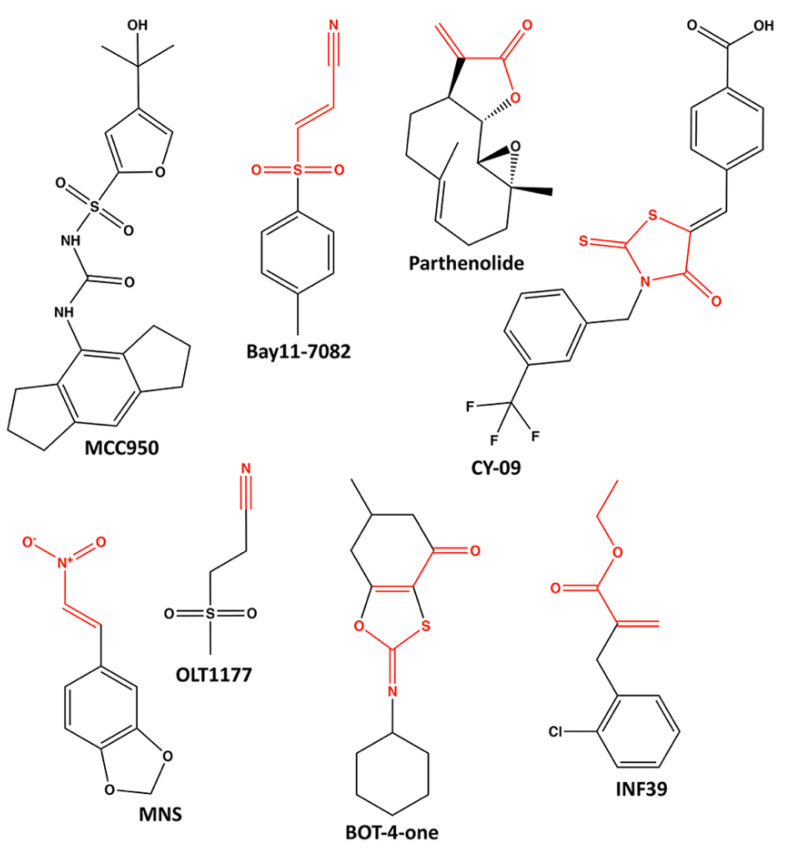
The prototypical model for NOD-like receptor (NLR) inflammasome assembly includes nucleotide-dependent activation of the NLR downstream of pathogen- or danger-associated molecular pattern (PAMP or DAMP) recognition, followed by nucleation of hetero-oligomeric platforms that lie upstream of inflammatory responses associated with innate immunity. As members of the STAND ATPases, the NLRs are generally thought to share a similar model of ATP-dependent activation and effect. However, recent observations have challenged this paradigm to reveal novel and complex biochemical processes to discern NLRs from other STAND proteins. In this review, we highlight past findings that identify the regulatory importance of conserved ATP-binding and hydrolysis motifs within the nucleotide-binding NACHT domain of NLRs and explore recent breakthroughs that generate connections between NLR protein structure and function. Indeed, newly deposited NLR structures for NLRC4 and NLRP3 have provided unique perspectives on the ATP-dependency of inflammasome activation. Novel molecular dynamic simulations of NLRP3 examined the active site of ADP- and ATP-bound models. The findings support distinctions in nucleotide-binding domain topology with occupancy of ATP or ADP that are in turn disseminated on to the global protein structure. Ultimately, studies continue to reveal how the ATP-binding and hydrolysis properties of NACHT domains in different NLRs integrate with signaling modules and binding partners to control innate immune responses at the molecular level.
Keywords: ATPase; NACHT domain; NLR; NLRP; NOD-like receptor; inflammasome; molecular dynamic simulation; nucleotide.
Conflict of interest statement
J.A.M. is cofounder and has an equity position in Arch Biopartners Inc. All other authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
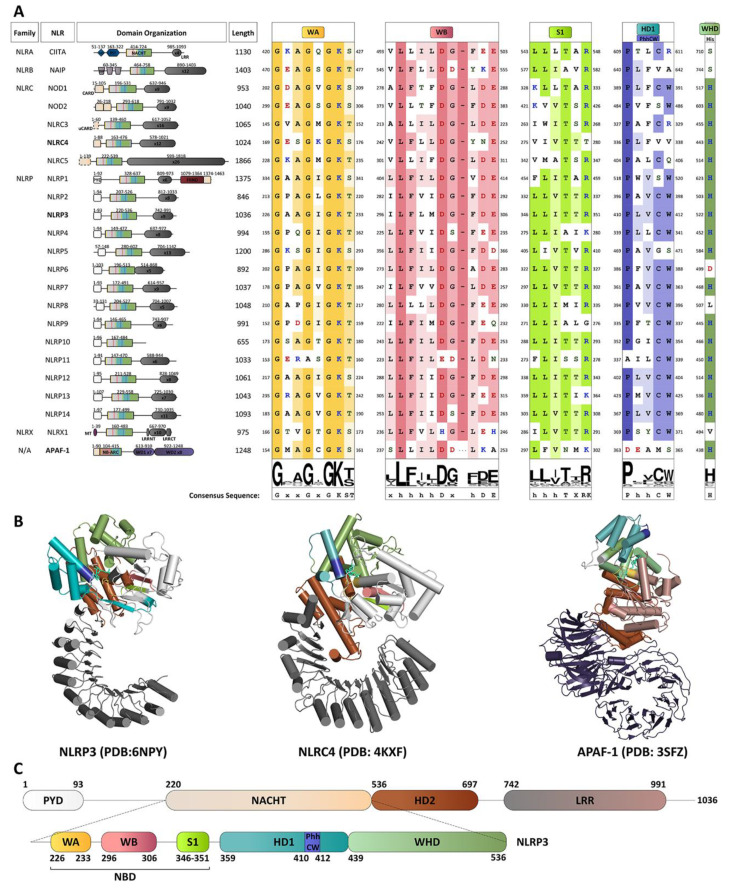
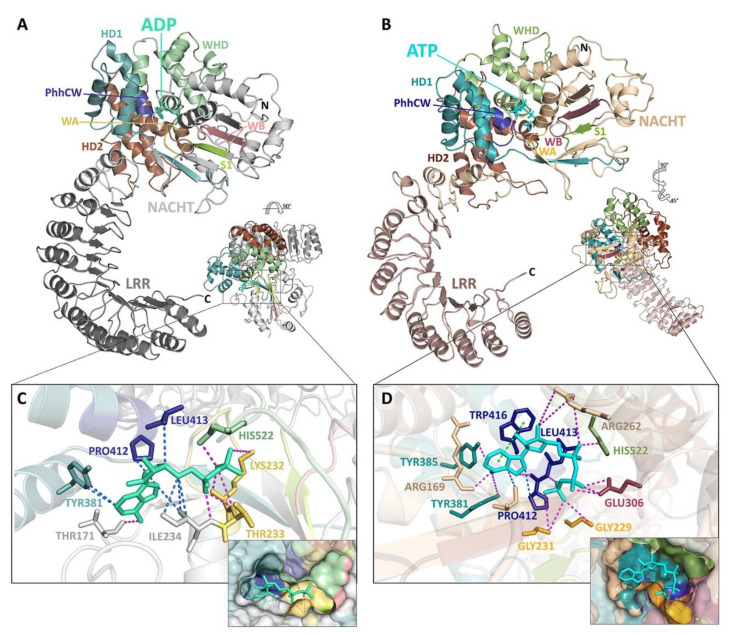
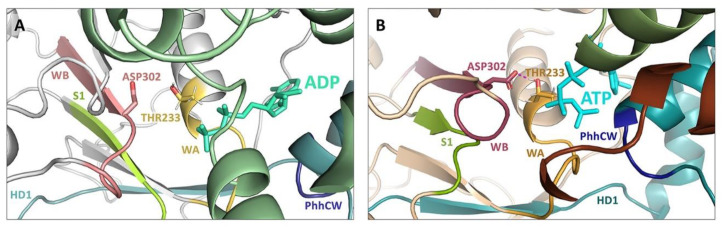
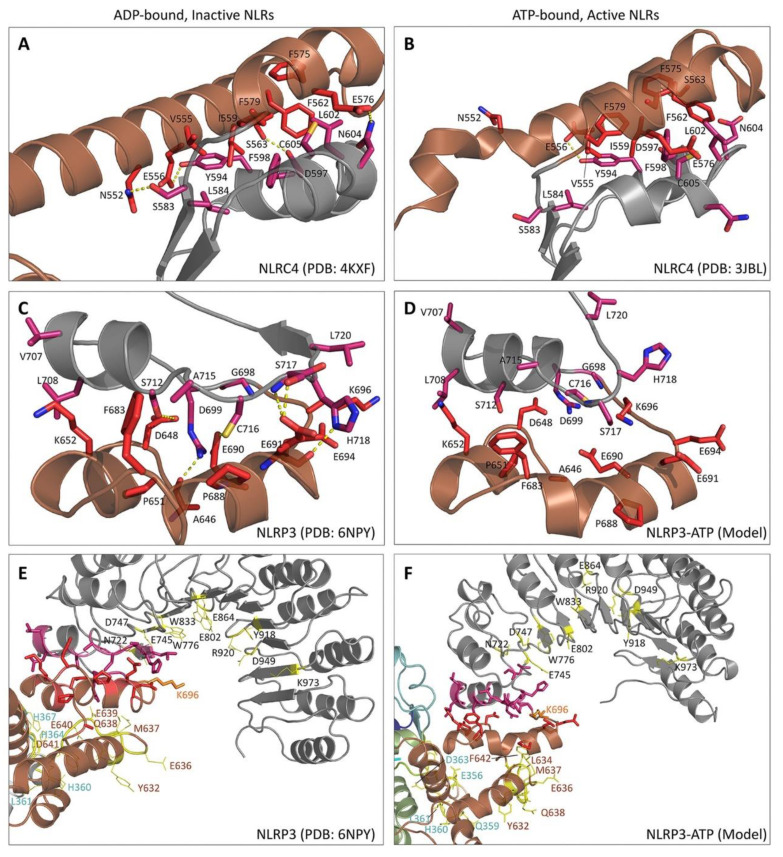
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
