

Cortical and striatal circuits together encode transitions in natural behavior
- PMID: 33036974
- PMCID: PMC11209674
- DOI: 10.1126/sciadv.abc1173
Cortical and striatal circuits together encode transitions in natural behavior
Abstract
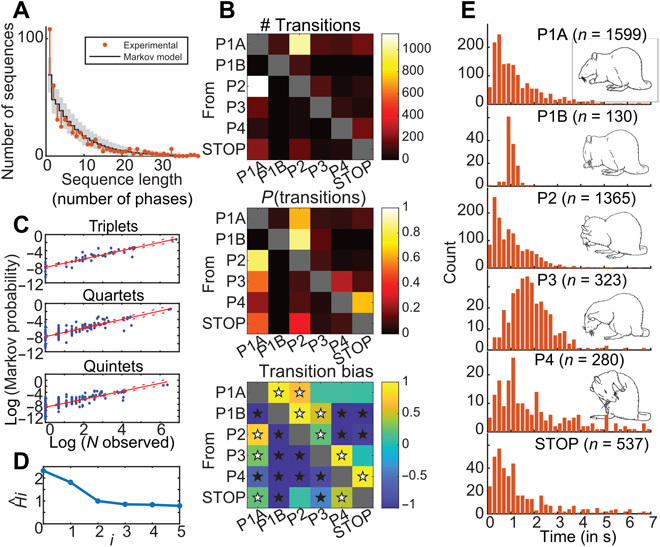
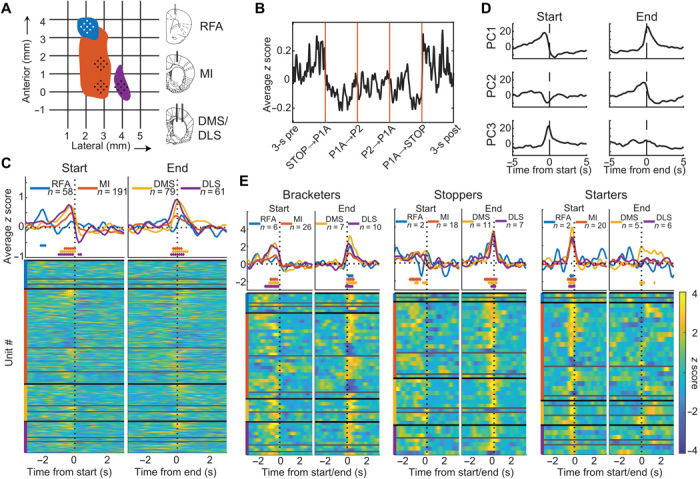
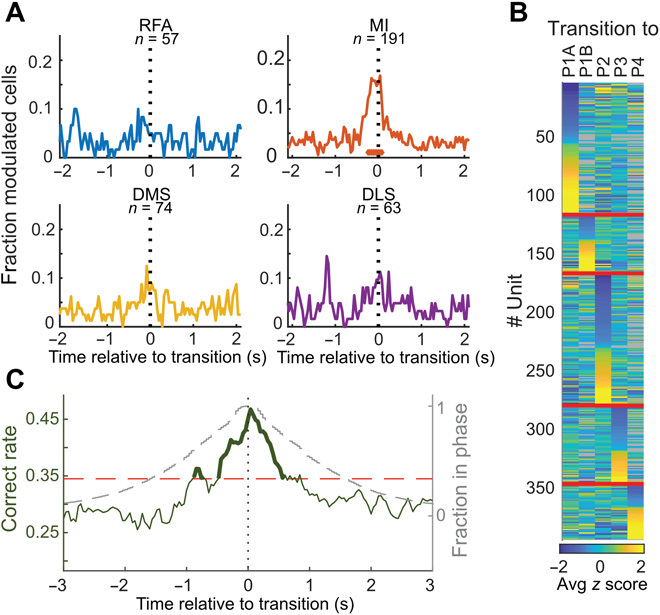
In natural behavior, we fluidly change from one type of activity to another in a sequence of motor actions. Corticostriatal circuits are thought to have a particularly important role in the construction of action sequences, but neuronal coding of a sequential behavior consisting of different motor programs has not been investigated at the circuit level in corticostriatal networks, making the exact nature of this involvement elusive. Here, we show, by analyzing spontaneous self-grooming in rats, that neuronal modulation in motor cortex and dorsal striatum is strongly related to transitions between behaviors. Our data suggest that longer action sequences in rodent grooming behavior emerge from stepwise control of individual behavioral transitions, where future actions are encoded differently depending on current motor state. This state-dependent motor coding was found to differentiate between rare behavioral transitions and as opposed to more habitual sequencing of actions.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures

Similar articles
-
Cortex, striatum and cerebellum: control of serial order in a grooming sequence.Exp Brain Res. 1992;90(2):275-90. doi: 10.1007/BF00227239. Exp Brain Res. 1992. PMID: 1397142
-
Inversely Active Striatal Projection Neurons and Interneurons Selectively Delimit Useful Behavioral Sequences.Curr Biol. 2018 Feb 19;28(4):560-573.e5. doi: 10.1016/j.cub.2018.01.031. Epub 2018 Feb 8. Curr Biol. 2018. PMID: 29429614 Free PMC article.
-
Corticostriatal dynamics encode the refinement of specific behavioral variability during skill learning.Elife. 2015 Sep 29;4:e09423. doi: 10.7554/eLife.09423. Elife. 2015. PMID: 26417950 Free PMC article.
-
Corticostriatal circuit mechanisms of value-based action selection: Implementation of reinforcement learning algorithms and beyond.Behav Brain Res. 2016 Sep 15;311:110-121. doi: 10.1016/j.bbr.2016.05.017. Epub 2016 May 9. Behav Brain Res. 2016. PMID: 27173430 Review.
-
Human and rodent homologies in action control: corticostriatal determinants of goal-directed and habitual action.Neuropsychopharmacology. 2010 Jan;35(1):48-69. doi: 10.1038/npp.2009.131. Neuropsychopharmacology. 2010. PMID: 19776734 Free PMC article. Review.
Cited by
-
Midbrain dopamine neurons arbiter OCD-like behavior.Proc Natl Acad Sci U S A. 2022 Nov 16;119(46):e2207545119. doi: 10.1073/pnas.2207545119. Epub 2022 Nov 7. Proc Natl Acad Sci U S A. 2022. PMID: 36343236 Free PMC article.
-
Control of striatal circuit development by the chromatin regulator Zswim6.Sci Adv. 2025 Jan 17;11(3):eadq6663. doi: 10.1126/sciadv.adq6663. Epub 2025 Jan 17. Sci Adv. 2025. PMID: 39823338 Free PMC article.
-
The basal ganglia control the detailed kinematics of learned motor skills.Nat Neurosci. 2021 Sep;24(9):1256-1269. doi: 10.1038/s41593-021-00889-3. Epub 2021 Jul 15. Nat Neurosci. 2021. PMID: 34267392 Free PMC article.
-
The neurobiology of parenting and infant-evoked aggression.Physiol Rev. 2025 Jan 1;105(1):315-381. doi: 10.1152/physrev.00036.2023. Epub 2024 Aug 15. Physiol Rev. 2025. PMID: 39146250 Free PMC article. Review.
-
Manipulation-specific cortical activity as mice handle food.Curr Biol. 2022 Nov 21;32(22):4842-4853.e6. doi: 10.1016/j.cub.2022.09.045. Epub 2022 Oct 14. Curr Biol. 2022. PMID: 36243014 Free PMC article.
References
-
- Mink J. W., The basal ganglia: Focused selection and inhibition of competing motor programs. Prog. Neurobiol. 50, 381–425 (1996). - PubMed
-
- Fujii N., Graybiel A. M., Representation of action sequence boundaries by macaque prefrontal cortical neurons. Science 301, 1246–1249 (2003). - PubMed
-
- Tanji J., Shima K., Role for supplementary motor area cells in planning several movements ahead. Nature 371, 413–416 (1994). - PubMed
-
- Barnes T. D., Kubota Y., Hu D., Jin D. Z., Graybiel A. M., Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature 437, 1158–1161 (2005). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
