

The reductive glycine pathway allows autotrophic growth of Desulfovibrio desulfuricans
- PMID: 33037220
- PMCID: PMC7547702
- DOI: 10.1038/s41467-020-18906-7
The reductive glycine pathway allows autotrophic growth of Desulfovibrio desulfuricans
Abstract
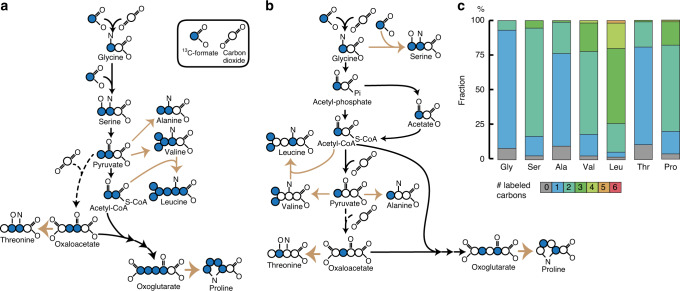
Six CO2 fixation pathways are known to operate in photoautotrophic and chemoautotrophic microorganisms. Here, we describe chemolithoautotrophic growth of the sulphate-reducing bacterium Desulfovibrio desulfuricans (strain G11) with hydrogen and sulphate as energy substrates. Genomic, transcriptomic, proteomic and metabolomic analyses reveal that D. desulfuricans assimilates CO2 via the reductive glycine pathway, a seventh CO2 fixation pathway. In this pathway, CO2 is first reduced to formate, which is reduced and condensed with a second CO2 to generate glycine. Glycine is further reduced in D. desulfuricans by glycine reductase to acetyl-P, and then to acetyl-CoA, which is condensed with another CO2 to form pyruvate. Ammonia is involved in the operation of the pathway, which is reflected in the dependence of the autotrophic growth rate on the ammonia concentration. Our study demonstrates microbial autotrophic growth fully supported by this highly ATP-efficient CO2 fixation pathway.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Smith, D. W. Ecological actions of sulfate-reducing bacteria. In The Sulfate-reducing Bacteria: Contemporary Perspectives (eds Odom, J.M. & Singleton, R.) 161–188 (Springer-Verlag, 1993).
-
- Jansen K, Thauer RK, Widdel F, Fuchs G. Carbon assimilation pathways in sulfate reducing bacteria. Formate, carbon dioxide, carbon monoxide, and acetate assimilation by Desulfovibrio baarsii. Arch. Microbiol. 1984;138:257–262. doi: 10.1007/BF00402132. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
