

Age-associated changes in the circulating human antibody repertoire are upregulated in autoimmunity
- PMID: 33042204
- PMCID: PMC7539520
- DOI: 10.1186/s12979-020-00193-x
Age-associated changes in the circulating human antibody repertoire are upregulated in autoimmunity
Abstract
Background: The immune system undergoes a myriad of changes with age. While it is known that antibody-secreting plasma and long-lived memory B cells change with age, it remains unclear how the binding profile of the circulating antibody repertoire is impacted.
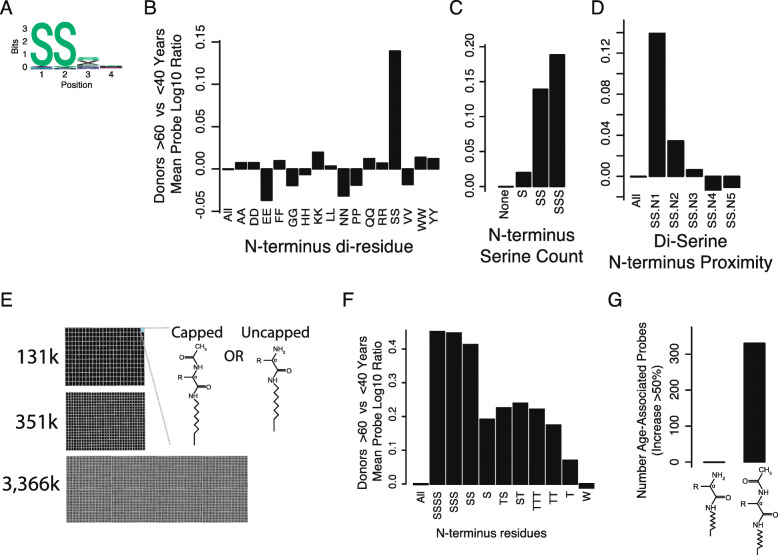
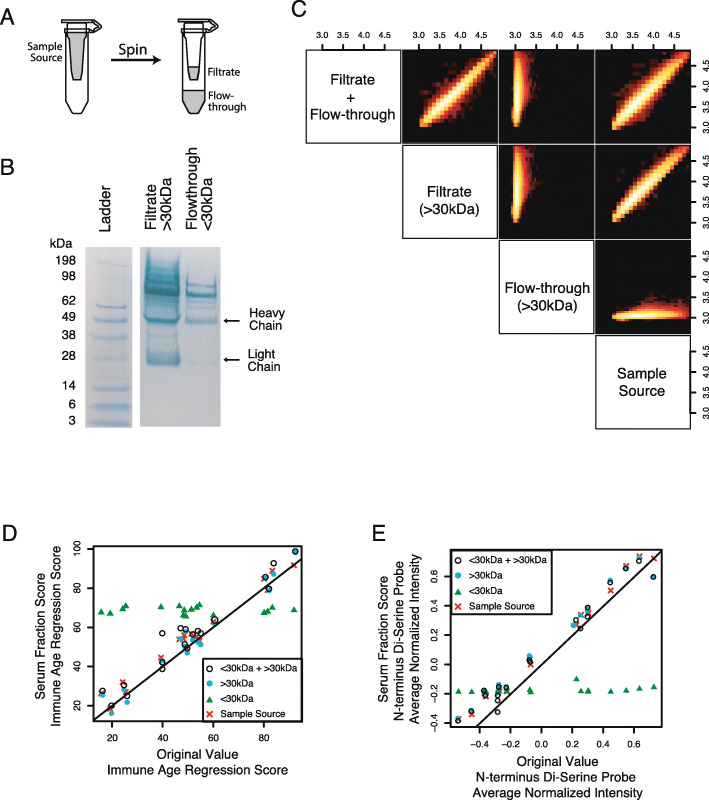
Results: To understand humoral immunity changes with respect to age, we characterized serum antibody binding to high density peptide microarrays in a diverse cohort of 1675 donors. We discovered thousands of peptides that bind antibodies in age-dependent fashion, many of which contain di-serine motifs. Peptide binding profiles were aggregated into an "immune age" by a machine learning regression model that was highly correlated with chronological age. Applying this regression model to previously-unobserved donors, we found that a donor's predicted immune age is longitudinally consistent over years, suggesting it could be a robust long-term biomarker of humoral immune ageing. Finally, we assayed serum from donors with autoimmune disease and found a significant association between "accelerated immune ageing" and autoimmune disease activity.
Conclusions: The circulating antibody repertoire has increased binding to thousands of di-serine peptide containing peptides in older donors, which can be represented as an immune age. Increased immune age is associated with autoimmune disease, acute inflammatory disease severity, and may be a broadly relevant biomarker of immune function in health, disease, and therapeutic intervention.
Keywords: Antibody binding profile; Antibody response; Auto-immune disease; Immune age; Immunosenescence; Machine learning; Peptide library.
© The Author(s) 2020.
Conflict of interest statement
Competing interestsAuthors have read the journals policy and the authors of this manuscript have the following competing interests: The authors are or were employed by HealthTell, Inc. and iCarbonX as indicated in the author’s affiliations. A patent application has been filed on the data presented. This does not alter the author’s adherence to the BMC Immunity and Ageing editorial policies.
Figures



References
LinkOut - more resources
Full Text Sources
