

Structural features and development of an assay platform of the parasite target deoxyhypusine synthase of Brugia malayi and Leishmania major
- PMID: 33044977
- PMCID: PMC7581365
- DOI: 10.1371/journal.pntd.0008762
Structural features and development of an assay platform of the parasite target deoxyhypusine synthase of Brugia malayi and Leishmania major
Abstract
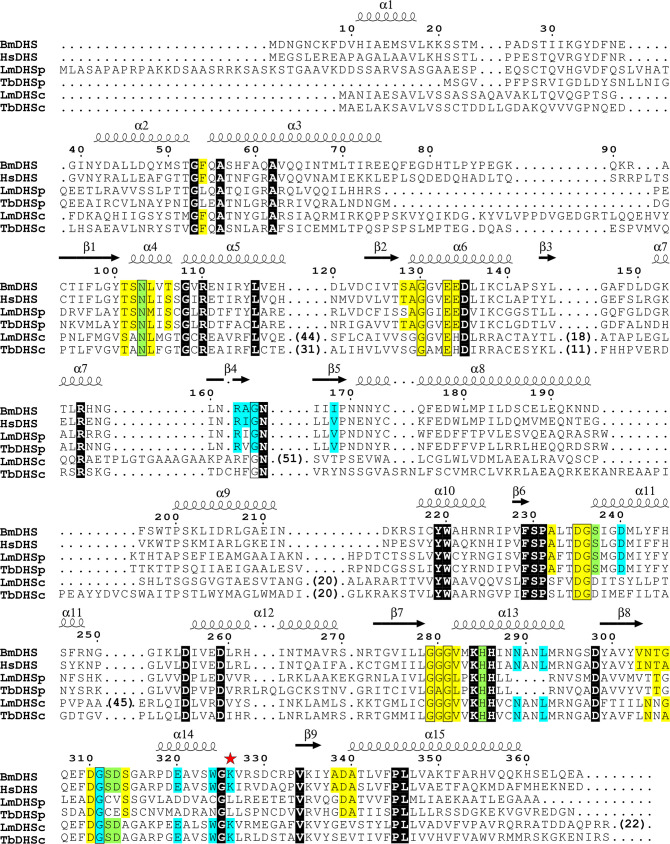
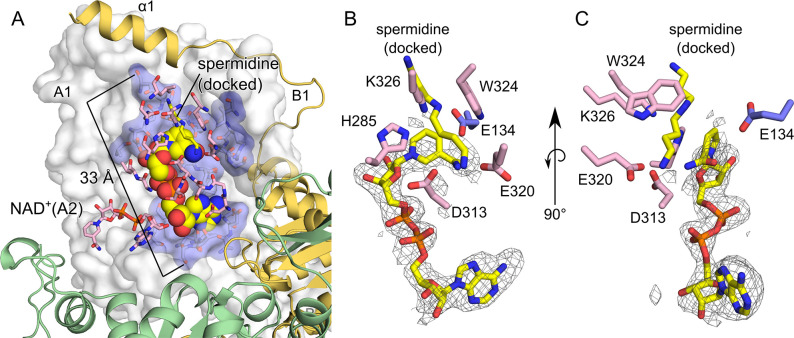
Deoxyhypusine synthase (DHS) catalyzes the first step of the post-translational modification of eukaryotic translation factor 5A (eIF5A), which is the only known protein containing the amino acid hypusine. Both proteins are essential for eukaryotic cell viability, and DHS has been suggested as a good candidate target for small molecule-based therapies against eukaryotic pathogens. In this work, we focused on the DHS enzymes from Brugia malayi and Leishmania major, the causative agents of lymphatic filariasis and cutaneous leishmaniasis, respectively. To enable B. malayi (Bm)DHS for future target-based drug discovery programs, we determined its crystal structure bound to cofactor NAD+. We also reported an in vitro biochemical assay for this enzyme that is amenable to a high-throughput screening format. The L. major genome encodes two DHS paralogs, and attempts to produce them recombinantly in bacterial cells were not successful. Nevertheless, we showed that ectopic expression of both LmDHS paralogs can rescue yeast cells lacking the endogenous DHS-encoding gene (dys1). Thus, functionally complemented dys1Δ yeast mutants can be used to screen for new inhibitors of the L. major enzyme. We used the known human DHS inhibitor GC7 to validate both in vitro and yeast-based DHS assays. Our results show that BmDHS is a homotetrameric enzyme that shares many features with its human homologue, whereas LmDHS paralogs are likely to form a heterotetrameric complex and have a distinct regulatory mechanism. We expect our work to facilitate the identification and development of new DHS inhibitors that can be used to validate these enzymes as vulnerable targets for therapeutic interventions against B. malayi and L. major infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
An experimental target-based platform in yeast for screening Plasmodium vivax deoxyhypusine synthase inhibitors.PLoS Negl Trop Dis. 2024 Dec 2;18(12):e0012690. doi: 10.1371/journal.pntd.0012690. eCollection 2024 Dec. PLoS Negl Trop Dis. 2024. PMID: 39621767 Free PMC article.
-
Yeast-Based High-Throughput Screens to Identify Novel Compounds Active against Brugia malayi.PLoS Negl Trop Dis. 2016 Jan 26;10(1):e0004401. doi: 10.1371/journal.pntd.0004401. eCollection 2016 Jan. PLoS Negl Trop Dis. 2016. PMID: 26812604 Free PMC article.
-
A new non-radioactive deoxyhypusine synthase assay adaptable to high throughput screening.Amino Acids. 2017 Nov;49(11):1793-1804. doi: 10.1007/s00726-017-2477-3. Epub 2017 Aug 17. Amino Acids. 2017. PMID: 28819816 Free PMC article.
-
Targeting polyamine metabolism for finding new drugs against leishmaniasis: a review.Mini Rev Med Chem. 2015;15(3):243-52. doi: 10.2174/138955751503150312141044. Mini Rev Med Chem. 2015. PMID: 25769972 Review.
-
Linking proteome and genome: how to identify parasite proteins.Trends Parasitol. 2001 Apr;17(4):198-202. doi: 10.1016/s1471-4922(00)01947-4. Trends Parasitol. 2001. PMID: 11282511 Review.
Cited by
-
Polyamine Metabolism for Drug Intervention in Trypanosomatids.Pathogens. 2024 Jan 16;13(1):79. doi: 10.3390/pathogens13010079. Pathogens. 2024. PMID: 38251386 Free PMC article. Review.
-
Development of an activity assay for characterizing deoxyhypusine synthase and its diverse reaction products.FEBS Open Bio. 2021 Jan;11(1):10-25. doi: 10.1002/2211-5463.13046. Epub 2020 Dec 8. FEBS Open Bio. 2021. PMID: 33247548 Free PMC article.
-
An experimental target-based platform in yeast for screening Plasmodium vivax deoxyhypusine synthase inhibitors.PLoS Negl Trop Dis. 2024 Dec 2;18(12):e0012690. doi: 10.1371/journal.pntd.0012690. eCollection 2024 Dec. PLoS Negl Trop Dis. 2024. PMID: 39621767 Free PMC article.
-
Antifungal activity of guanidine compounds.Braz J Microbiol. 2025 Jun;56(2):1049-1059. doi: 10.1007/s42770-025-01625-w. Epub 2025 Feb 12. Braz J Microbiol. 2025. PMID: 39934527
-
The eukaryotic initiation factor 5A (eIF5A1), the molecule, mechanisms and recent insights into the pathophysiological roles.Cell Biosci. 2021 Dec 24;11(1):219. doi: 10.1186/s13578-021-00733-y. Cell Biosci. 2021. PMID: 34952646 Free PMC article. Review.
References
-
- Klohe K, Amuasi J, Kaducu JM, Haavardsson I, Bogatyreva E, Onarheim KH, et al. The 2017 Oslo conference report on neglected tropical diseases and emerging/re-emerging infectious diseases—Focus on populations underserved. Infect Dis Poverty. 2019;8: 1–10. 10.1186/s40249-018-0513-5 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
