

Balancing the Opposing Principles That Govern Peroxisome Homeostasis
- PMID: 33046344
- PMCID: PMC7880872
- DOI: 10.1016/j.tibs.2020.09.006
Balancing the Opposing Principles That Govern Peroxisome Homeostasis
Abstract
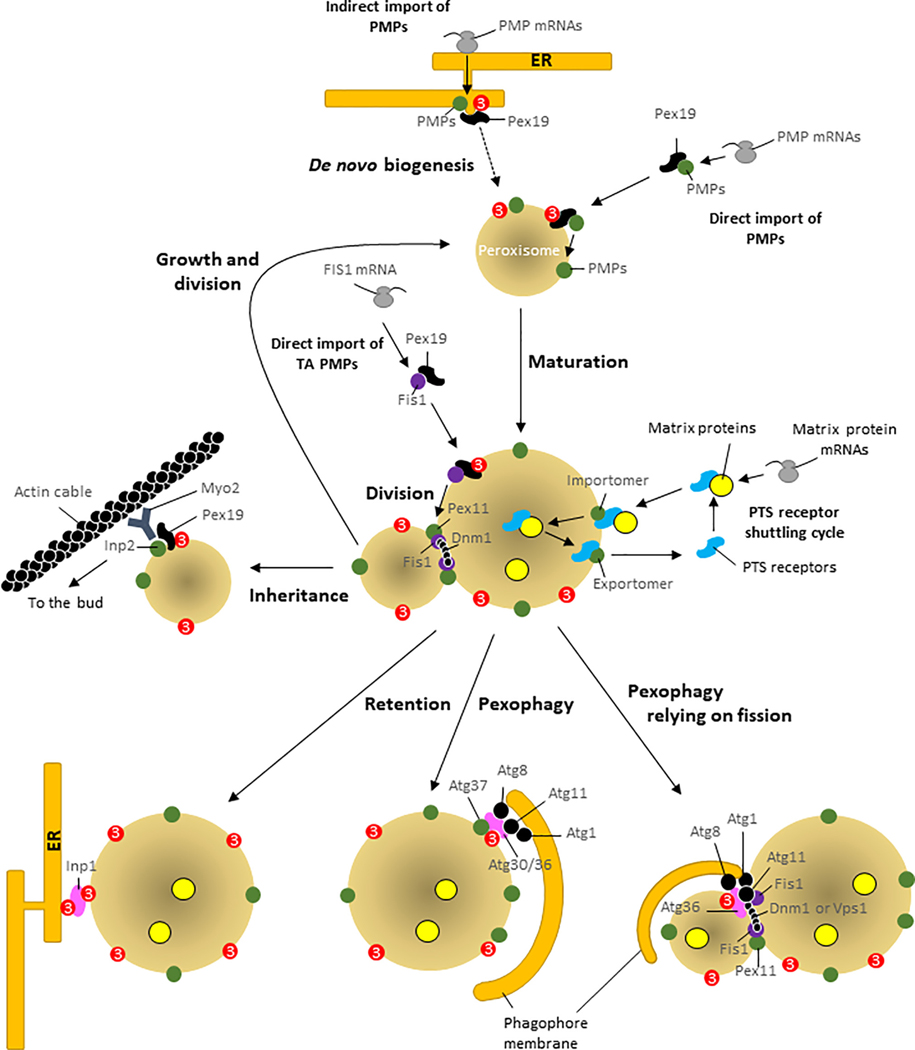
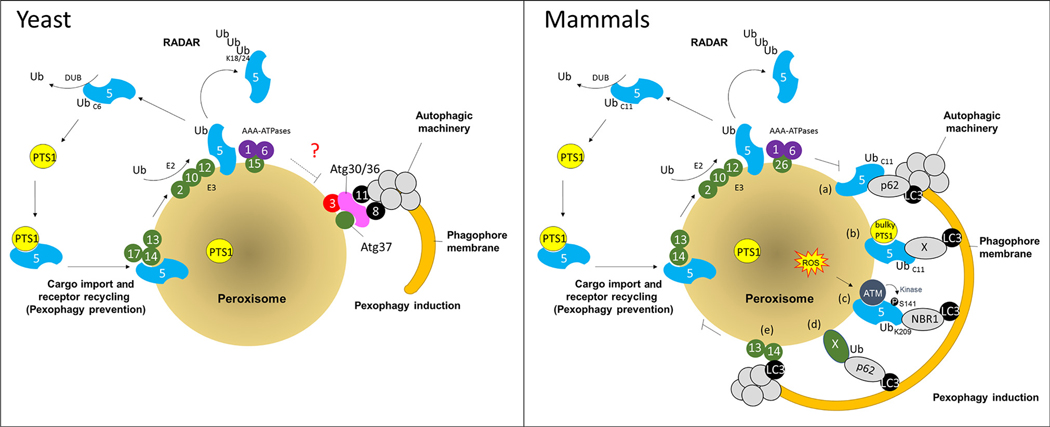
Despite major advances in our understanding of players and mechanisms involved in peroxisome biogenesis and peroxisome degradation, very few studies have focused on unraveling the multi-layered connections between, and the coordination of, these two opposing processes that regulate peroxisome homeostasis. The intersection between these processes also provides exciting avenues for future research. This review highlights the links between peroxisome biogenesis and degradation, incorporating an integrative approach that is critical not only for a mechanistic understanding, but also for manipulating the balance between these processes in relevant disease models.
Keywords: crosstalk; homeostasis; peroxisome biogenesis; peroxisome disorders; pexophagy.
Copyright © 2020 Elsevier Ltd. All rights reserved.
Figures

References
-
- Meinecke M et al. (2010) The peroxisomal importomer constitutes a large and highly dynamic pore. Nat. Cell Biol. 12, 273–277 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
