

Swine acute diarrhea syndrome coronavirus replication in primary human cells reveals potential susceptibility to infection
- PMID: 33046644
- PMCID: PMC7604506
- DOI: 10.1073/pnas.2001046117
Swine acute diarrhea syndrome coronavirus replication in primary human cells reveals potential susceptibility to infection
Abstract
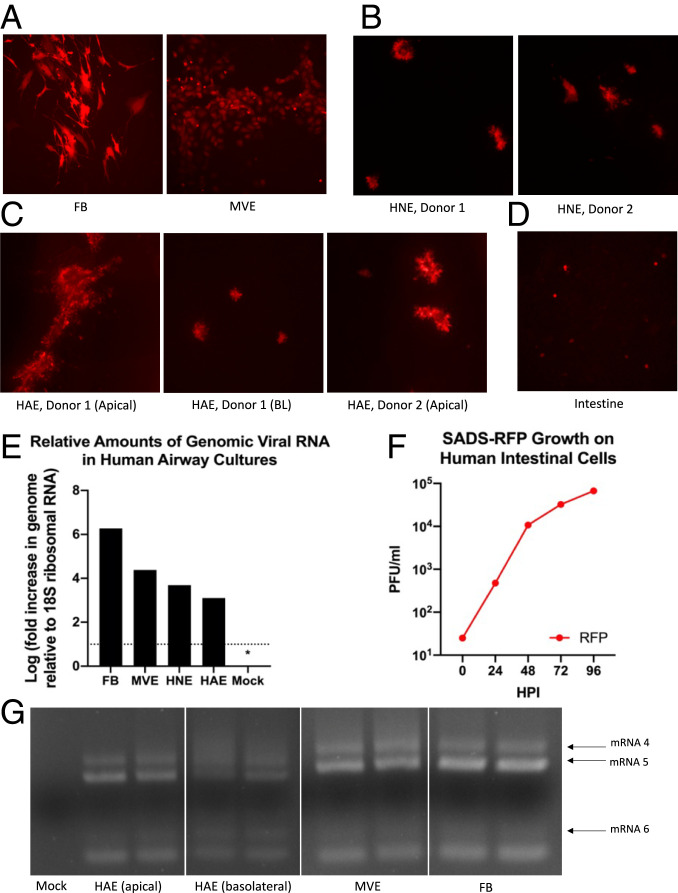
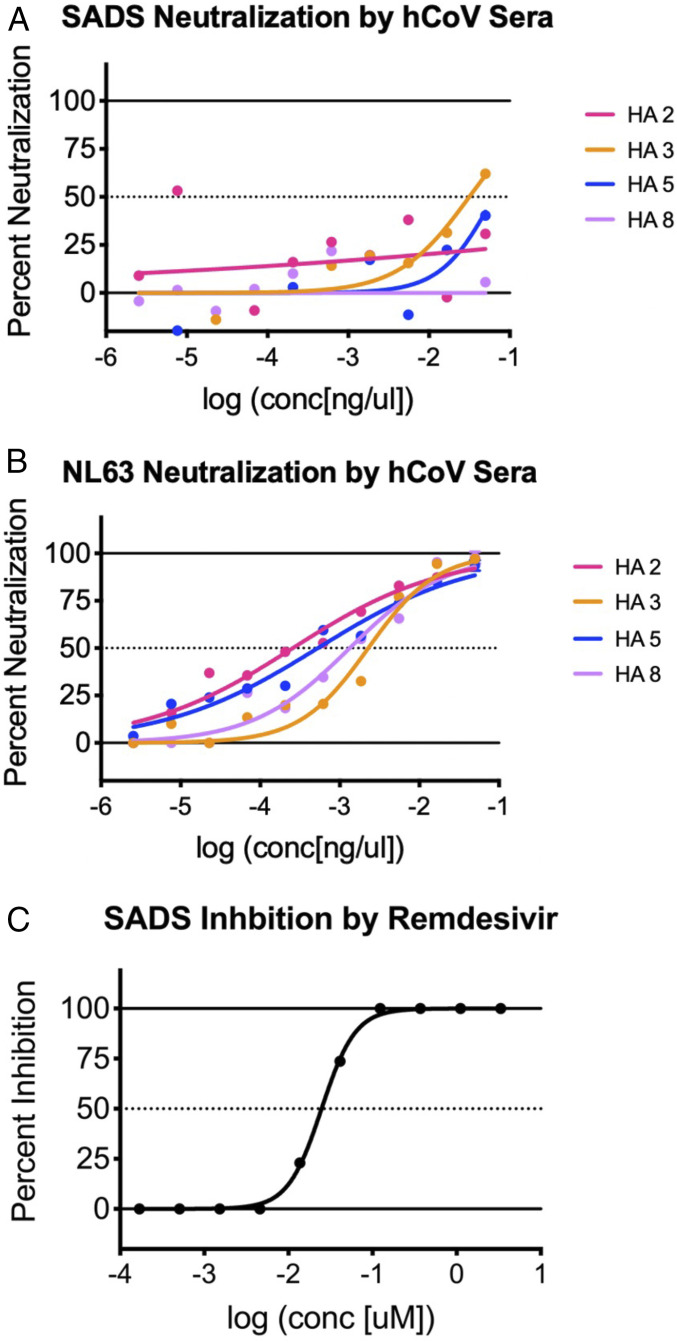
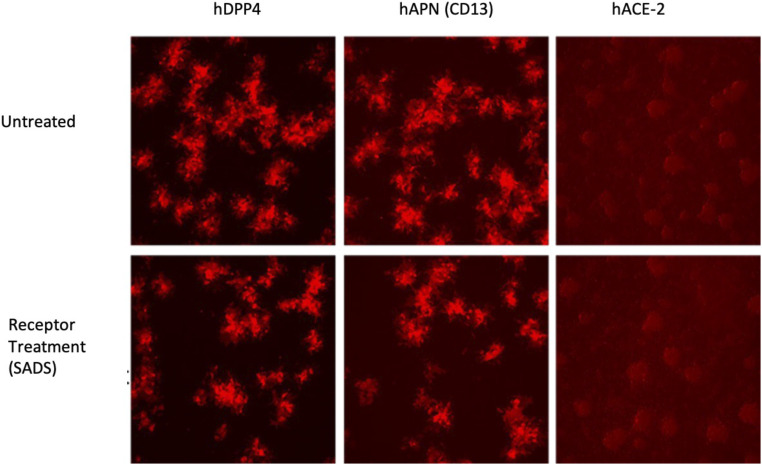
Zoonotic coronaviruses represent an ongoing threat, yet the myriads of circulating animal viruses complicate the identification of higher-risk isolates that threaten human health. Swine acute diarrhea syndrome coronavirus (SADS-CoV) is a newly discovered, highly pathogenic virus that likely evolved from closely related HKU2 bat coronaviruses, circulating in Rhinolophus spp. bats in China and elsewhere. As coronaviruses cause severe economic losses in the pork industry and swine are key intermediate hosts of human disease outbreaks, we synthetically resurrected a recombinant virus (rSADS-CoV) as well as a derivative encoding tomato red fluorescent protein (tRFP) in place of ORF3. rSADS-CoV replicated efficiently in a variety of continuous animal and primate cell lines, including human liver and rectal carcinoma cell lines. Of concern, rSADS-CoV also replicated efficiently in several different primary human lung cell types, as well as primary human intestinal cells. rSADS-CoV did not use human coronavirus ACE-2, DPP4, or CD13 receptors for docking and entry. Contemporary human donor sera neutralized the group I human coronavirus NL63, but not rSADS-CoV, suggesting limited human group I coronavirus cross protective herd immunity. Importantly, remdesivir, a broad-spectrum nucleoside analog that is effective against other group 1 and 2 coronaviruses, efficiently blocked rSADS-CoV replication in vitro. rSADS-CoV demonstrated little, if any, replicative capacity in either immune-competent or immunodeficient mice, indicating a critical need for improved animal models. Efficient growth in primary human lung and intestinal cells implicate SADS-CoV as a potential higher-risk emerging coronavirus pathogen that could negatively impact the global economy and human health.
Keywords: One Health; SADS; coronavirus; emerging infectious disease.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
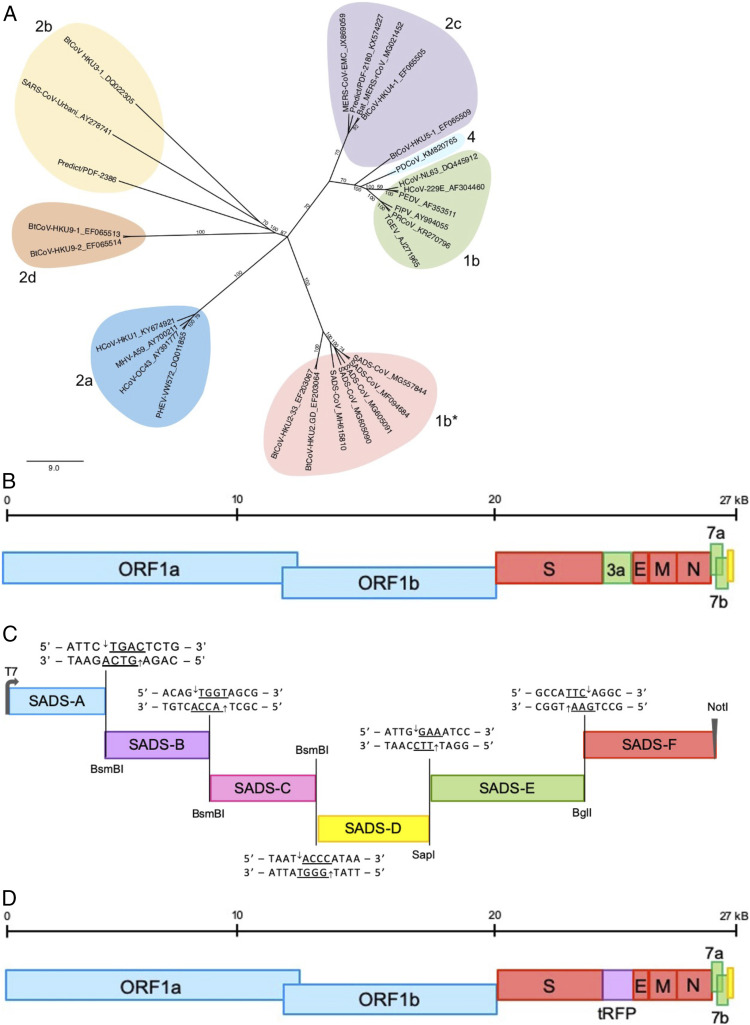
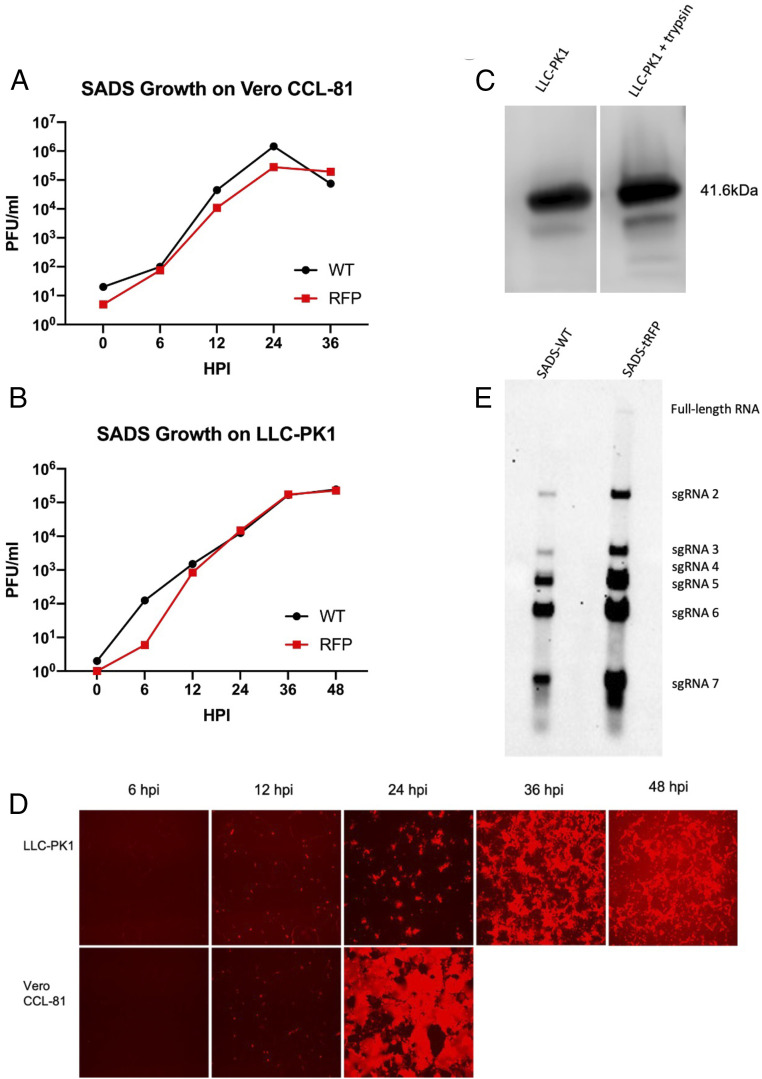
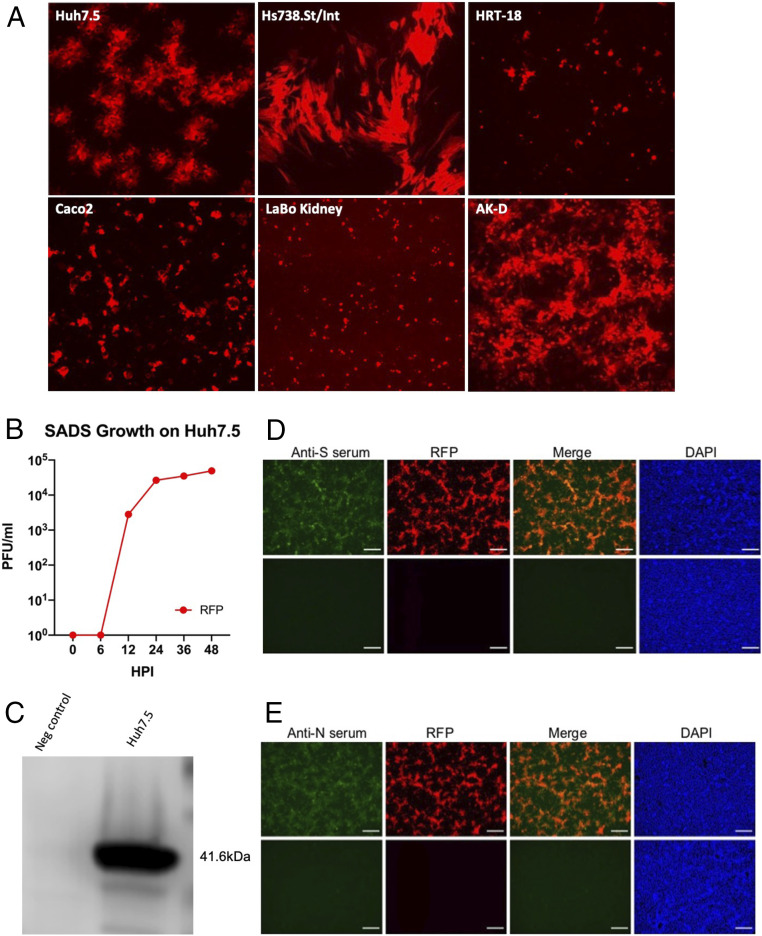
Comment in
-
Viruses from poultry and livestock pose continuous threats to human beings.Proc Natl Acad Sci U S A. 2021 Jan 19;118(3):e2022344118. doi: 10.1073/pnas.2022344118. Proc Natl Acad Sci U S A. 2021. PMID: 33414272 Free PMC article. No abstract available.
References
-
- WHO , Middle East respiratory syndrome coronavirus (MERS-CoV)—Update, https://www.who.int/csr/don/2014_07_23_mers/en/. Accessed 7 October 2019.
-
- WHO , Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003, https://www.who.int/csr/sars/country/table2004_04_21/en/. Accessed 7 October 2019.
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
