

The structure of a calsequestrin filament reveals mechanisms of familial arrhythmia
- PMID: 33046906
- PMCID: PMC7718342
- DOI: 10.1038/s41594-020-0510-9
The structure of a calsequestrin filament reveals mechanisms of familial arrhythmia
Abstract
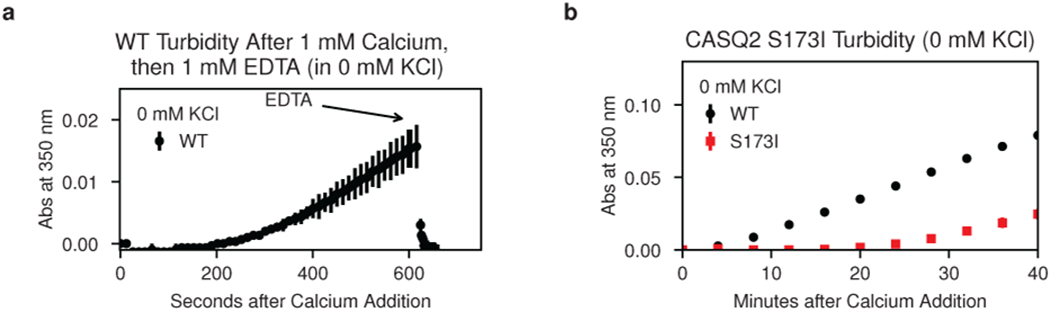
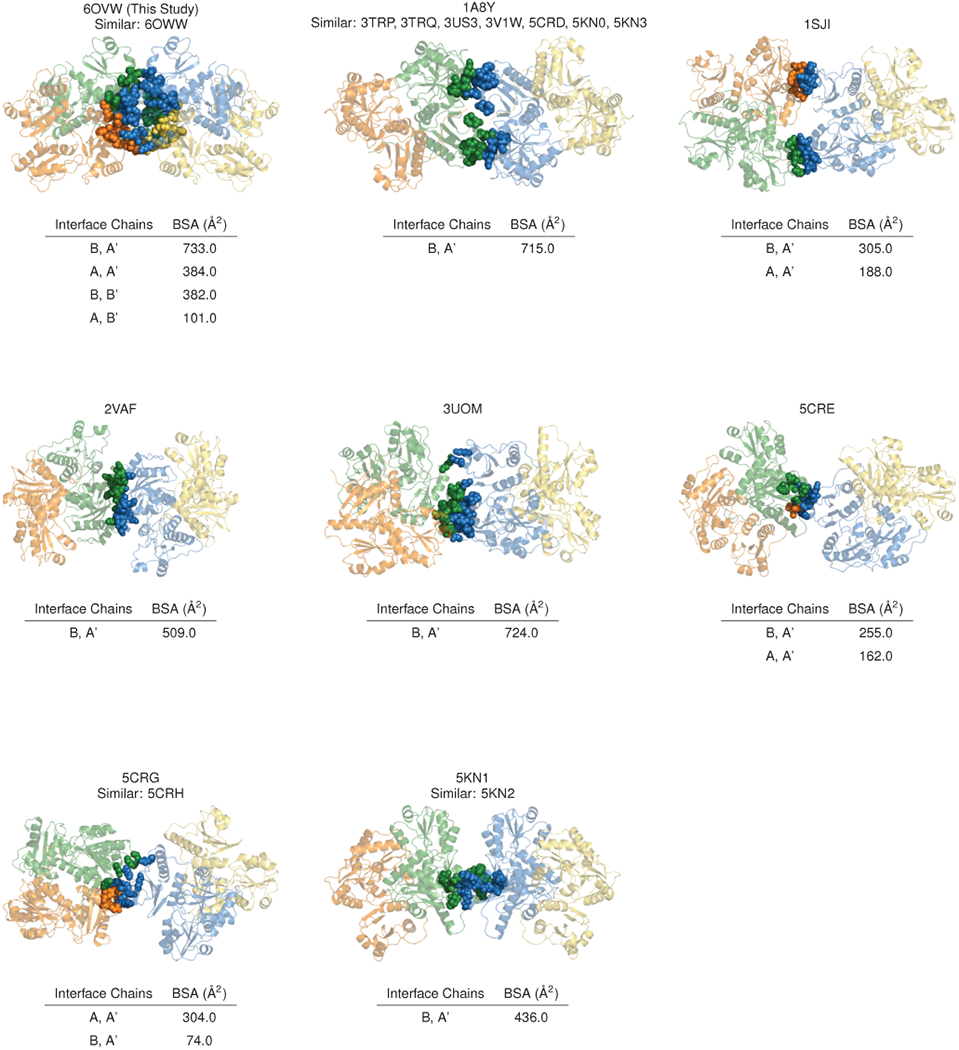
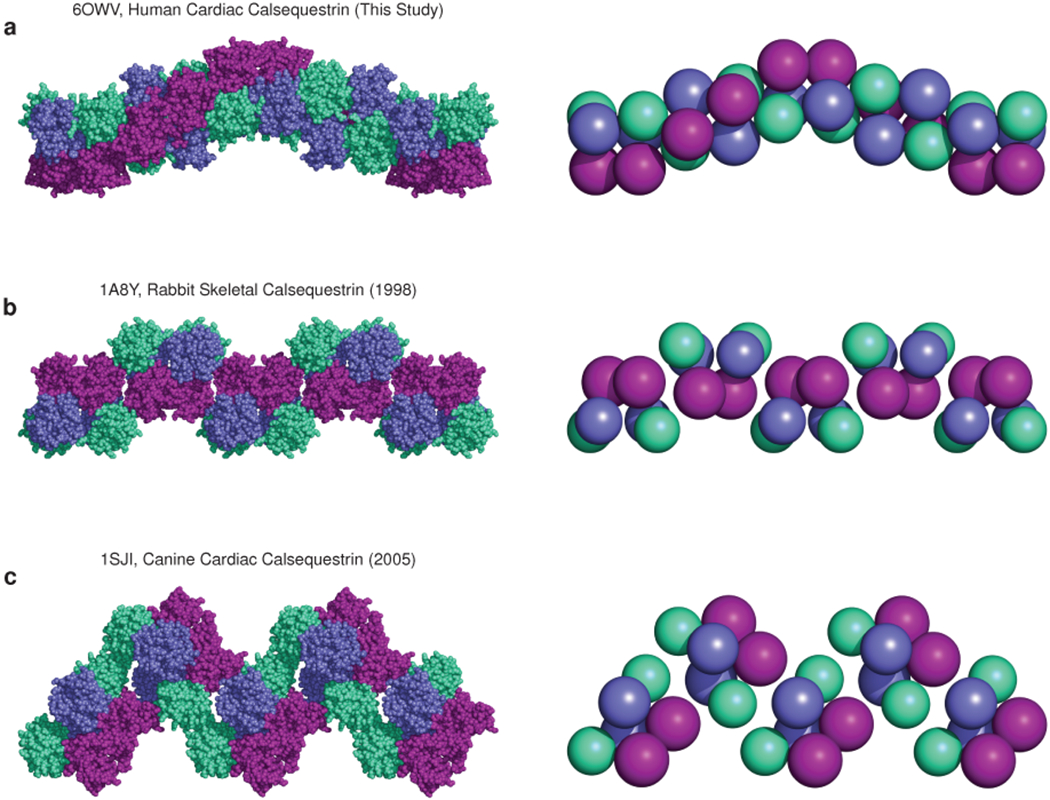
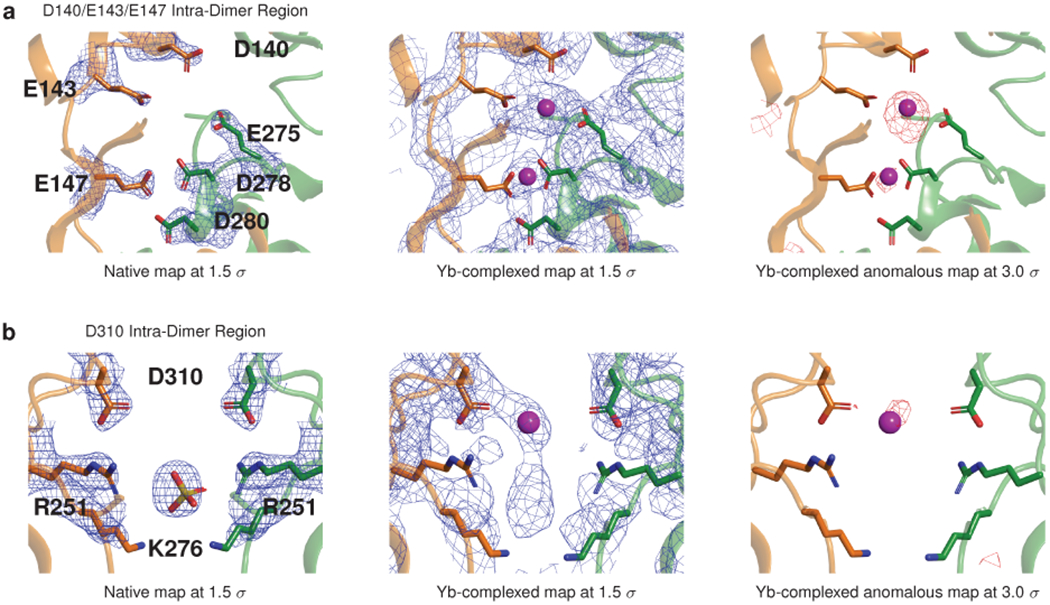
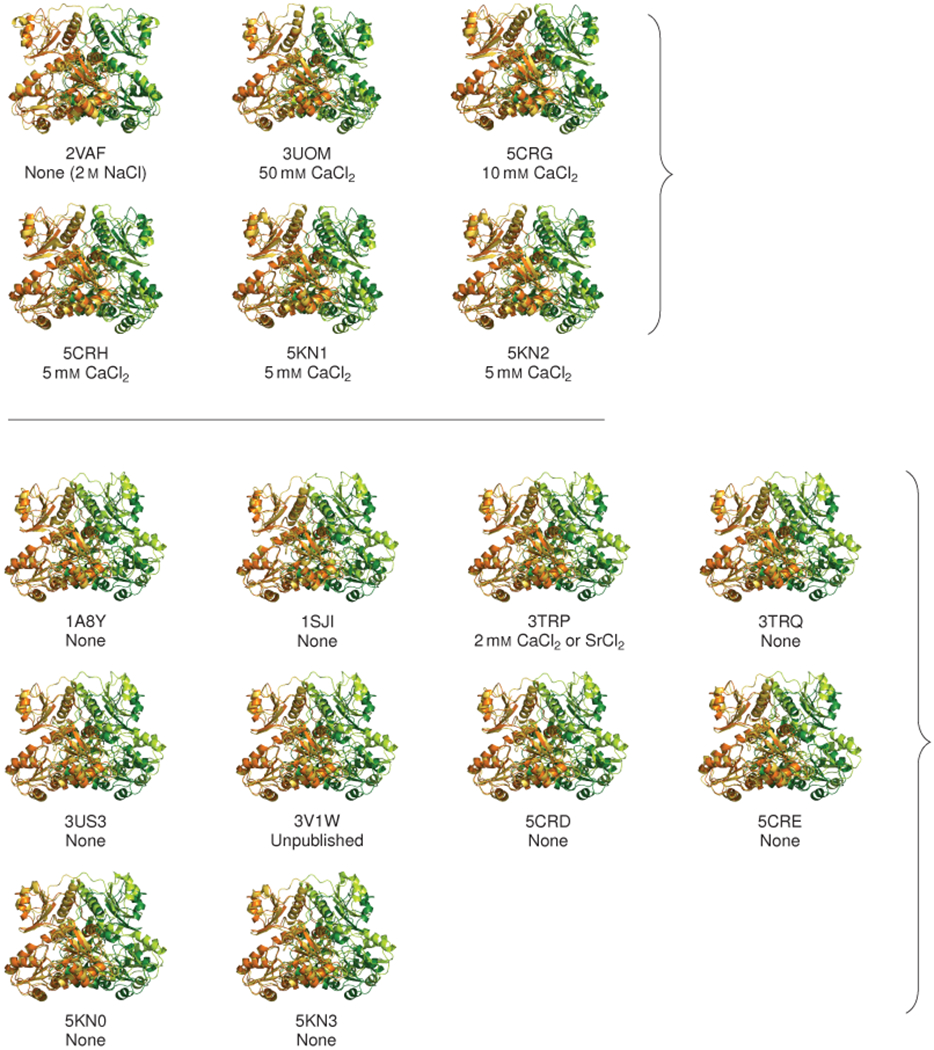
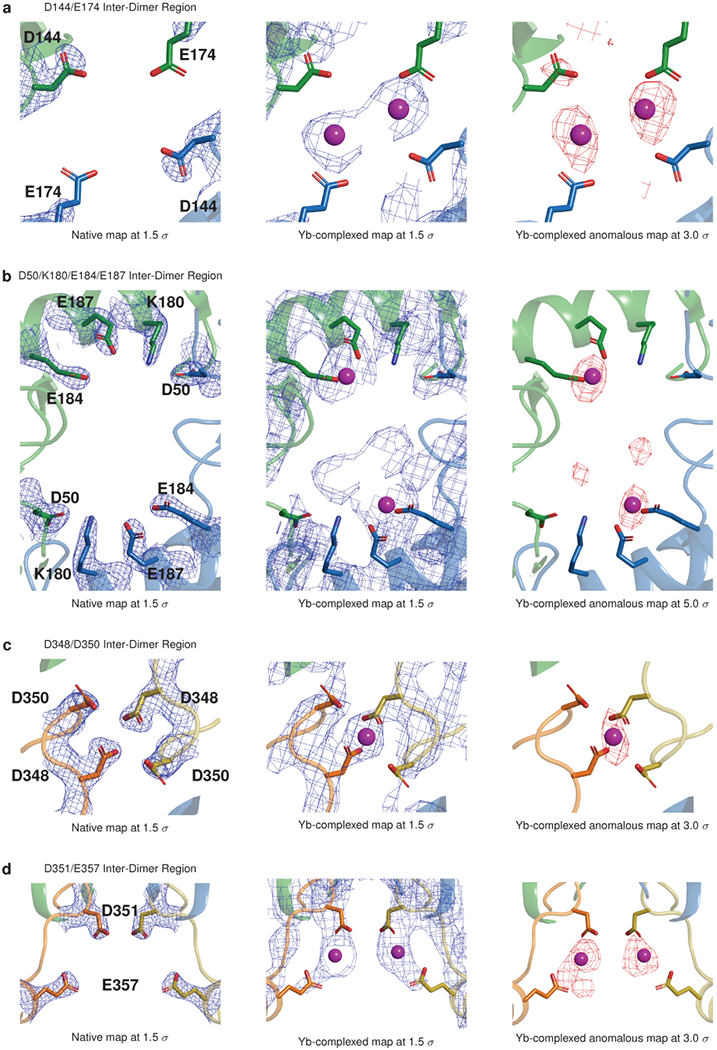
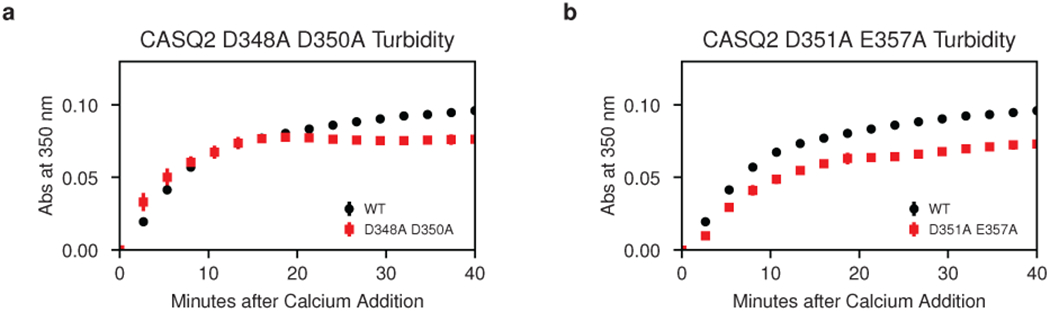
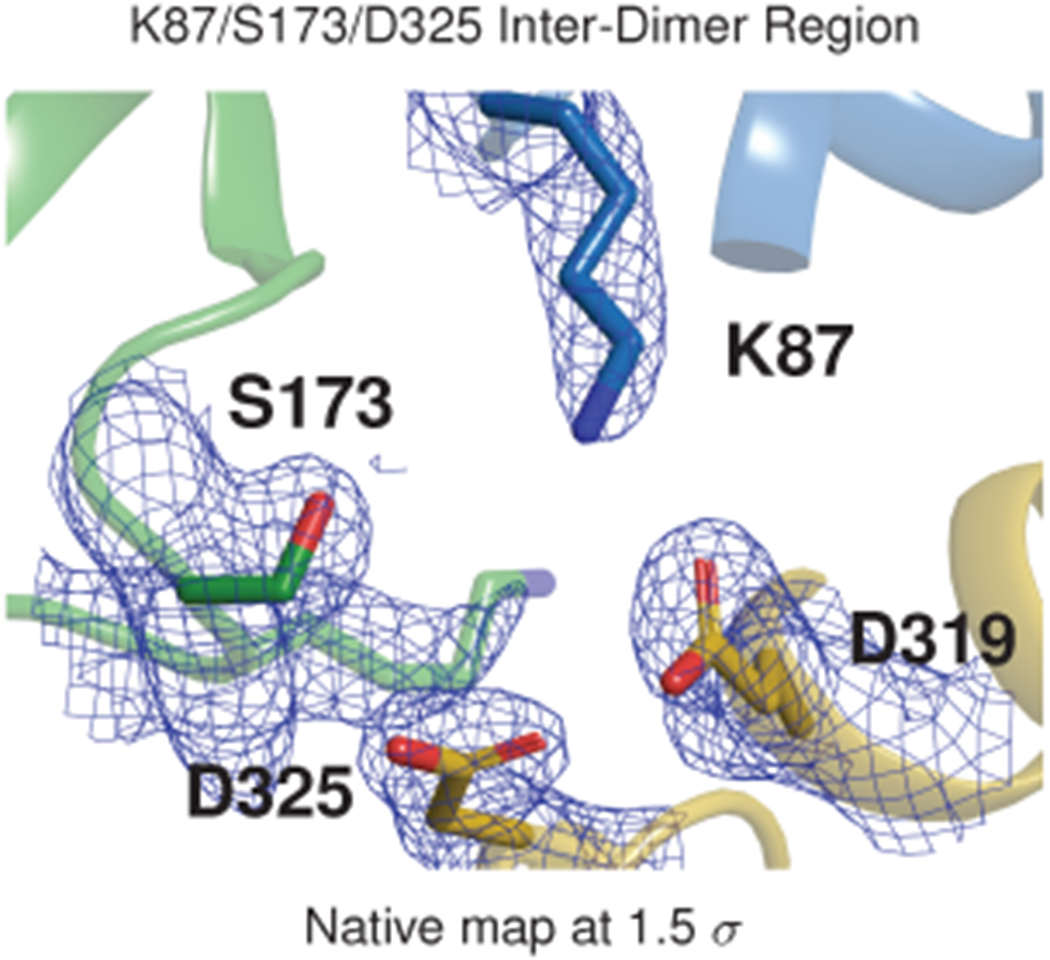
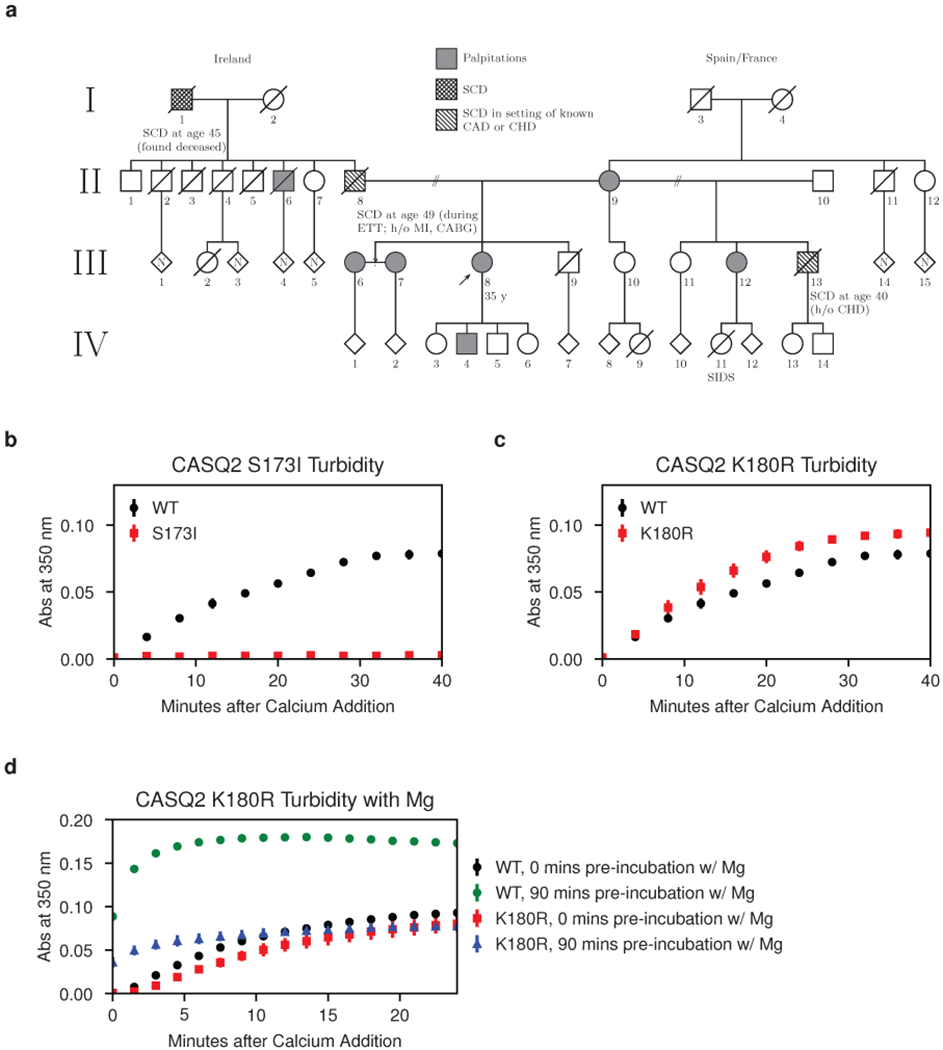
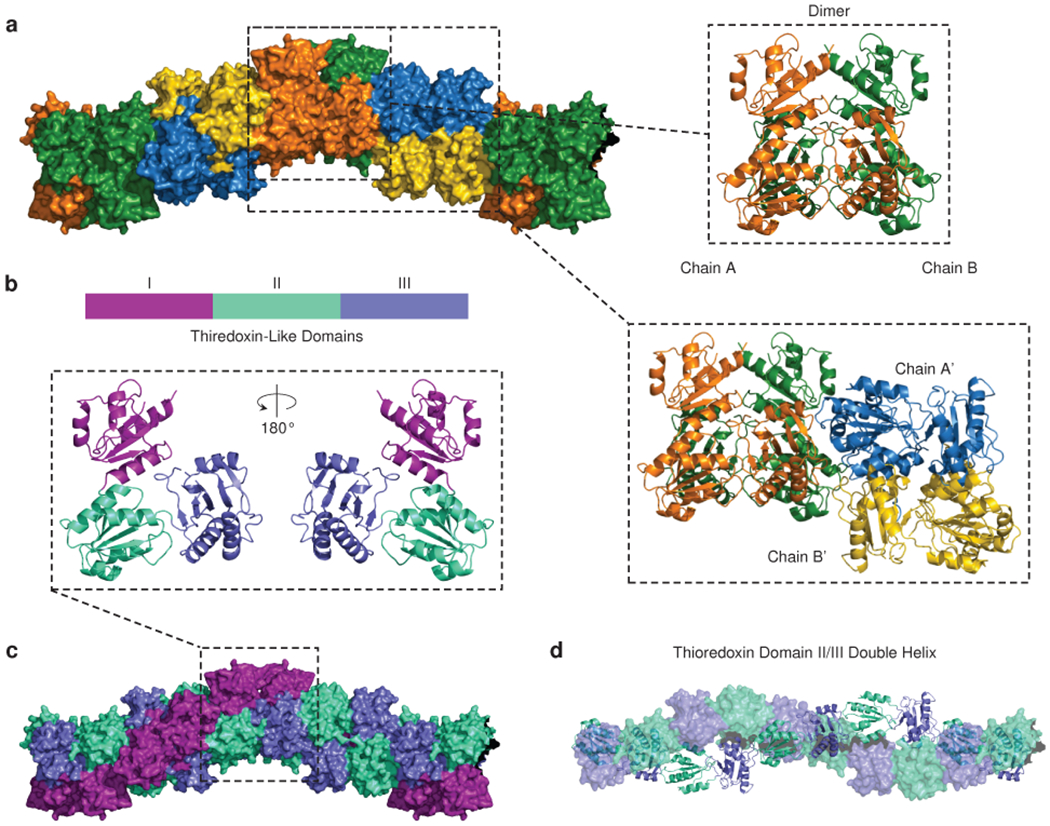
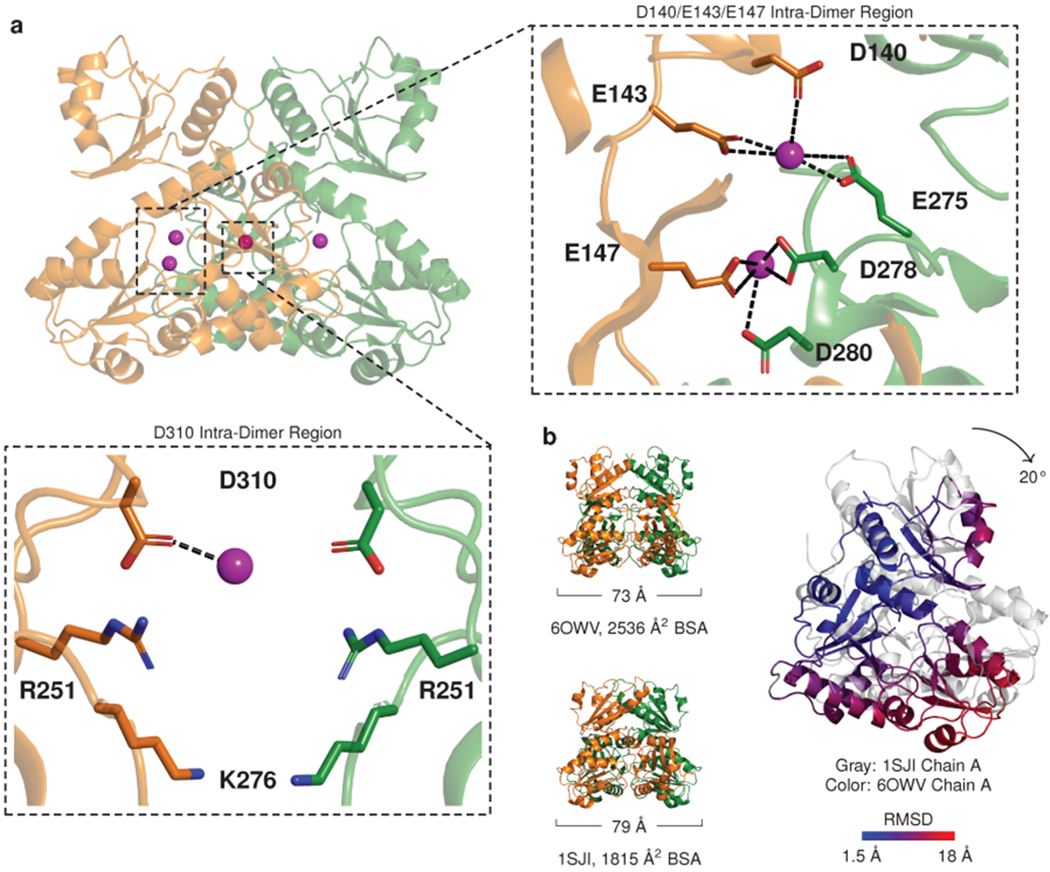
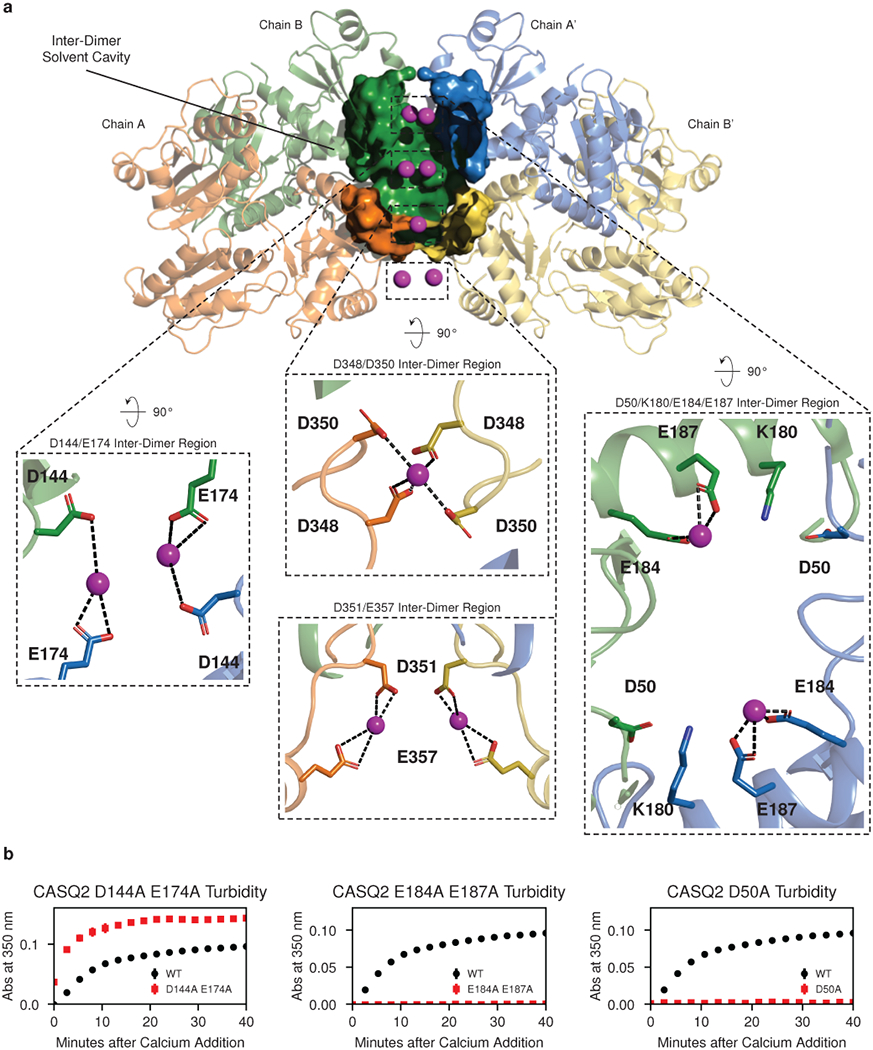
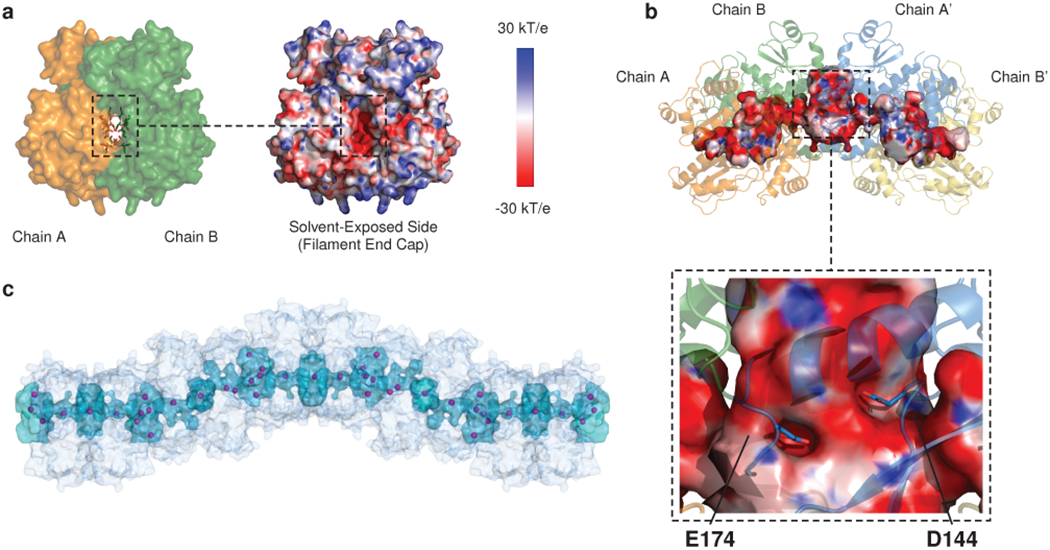
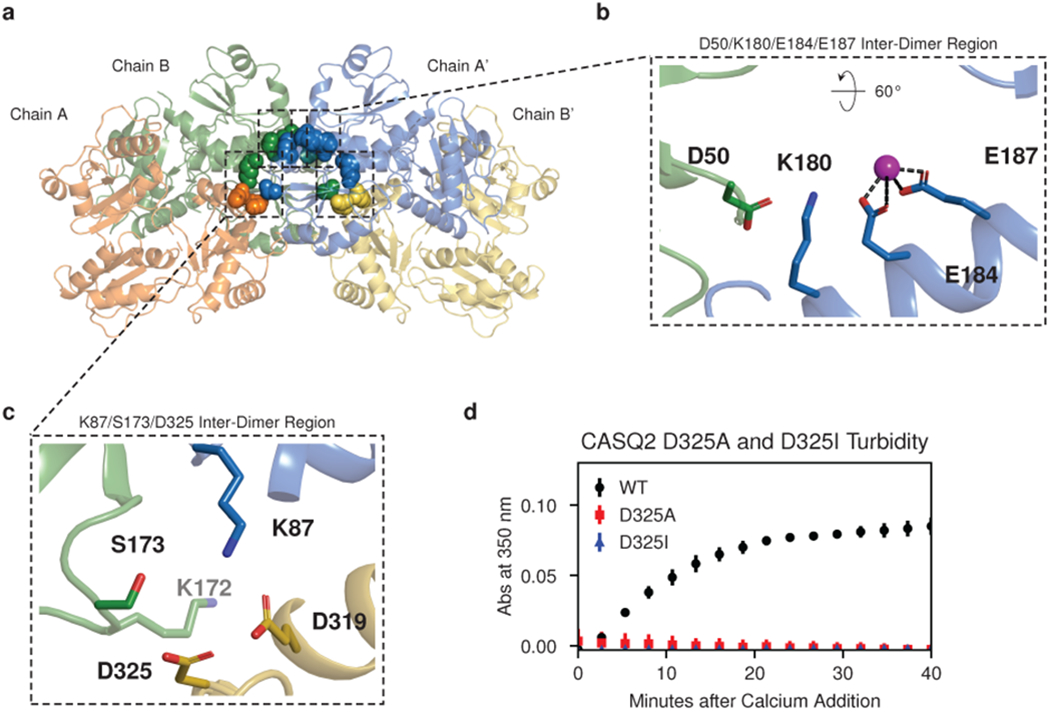
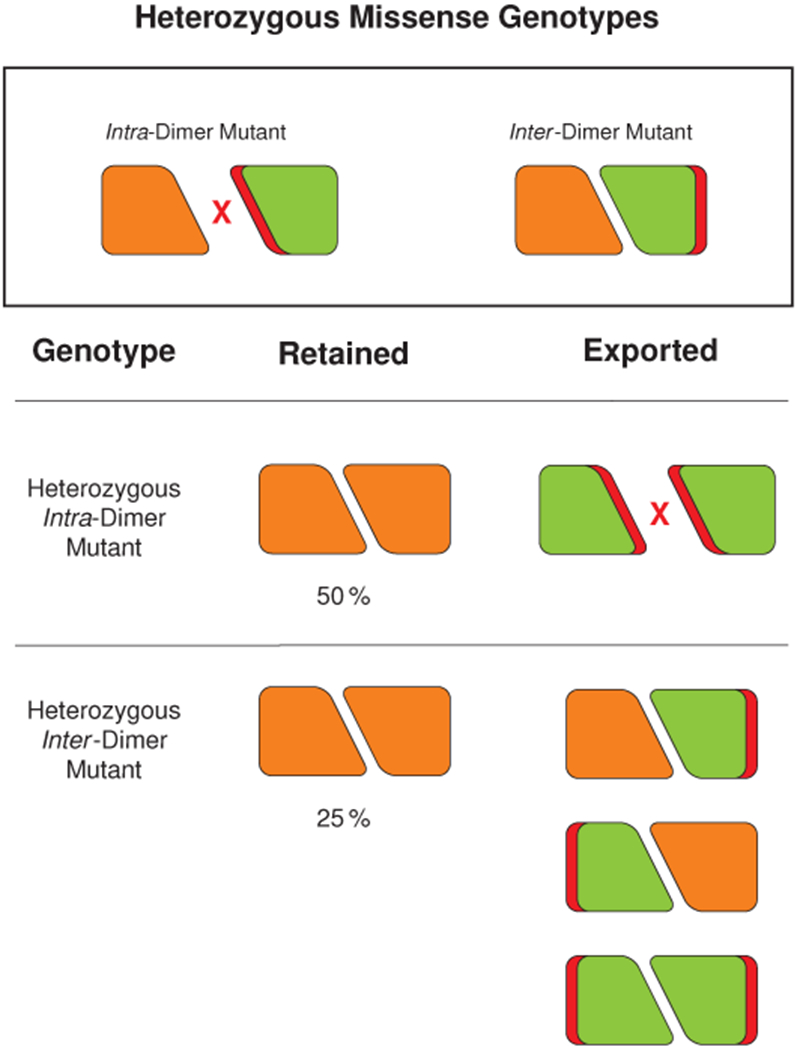
Mutations in the calcium-binding protein calsequestrin cause the highly lethal familial arrhythmia catecholaminergic polymorphic ventricular tachycardia (CPVT). In vivo, calsequestrin multimerizes into filaments, but there is not yet an atomic-resolution structure of a calsequestrin filament. We report a crystal structure of a human cardiac calsequestrin filament with supporting mutational analysis and in vitro filamentation assays. We identify and characterize a new disease-associated calsequestrin mutation, S173I, that is located at the filament-forming interface, and further show that a previously reported dominant disease mutation, K180R, maps to the same surface. Both mutations disrupt filamentation, suggesting that disease pathology is due to defects in multimer formation. An ytterbium-derivatized structure pinpoints multiple credible calcium sites at filament-forming interfaces, explaining the atomic basis of calsequestrin filamentation in the presence of calcium. Our study thus provides a unifying molecular mechanism through which dominant-acting calsequestrin mutations provoke lethal arrhythmias.
Conflict of interest statement
Declaration of Interests
The authors declare no competing interests.
Figures

References
-
- Bers DM Macromolecular complexes regulating cardiac ryanodine receptor function. J. Mol. Cell. Cardiol 37, 417–429 (2004). - PubMed
-
- Guerrero-Hernández A et al. in Calcium Signaling (ed Islam MS) 337–370 (Springer International Publishing, 2020).
-
- MacLennan DH Isolation of a second form of calsequestrin. J. Biol. Chem 249, 980–984 (1974). - PubMed
Methods References
-
- Winter G xia2: an expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr 43, 186–190 (2010).
-
- Evans P Scaling and assessment of data quality. Acta Crystallogr. D Biol. Crystallogr 62, 72–82 (2006). - PubMed
