

Hydroxylated sphingolipid biosynthesis regulates photoreceptor apical domain morphogenesis
- PMID: 33048164
- PMCID: PMC7557679
- DOI: 10.1083/jcb.201911100
Hydroxylated sphingolipid biosynthesis regulates photoreceptor apical domain morphogenesis
Abstract
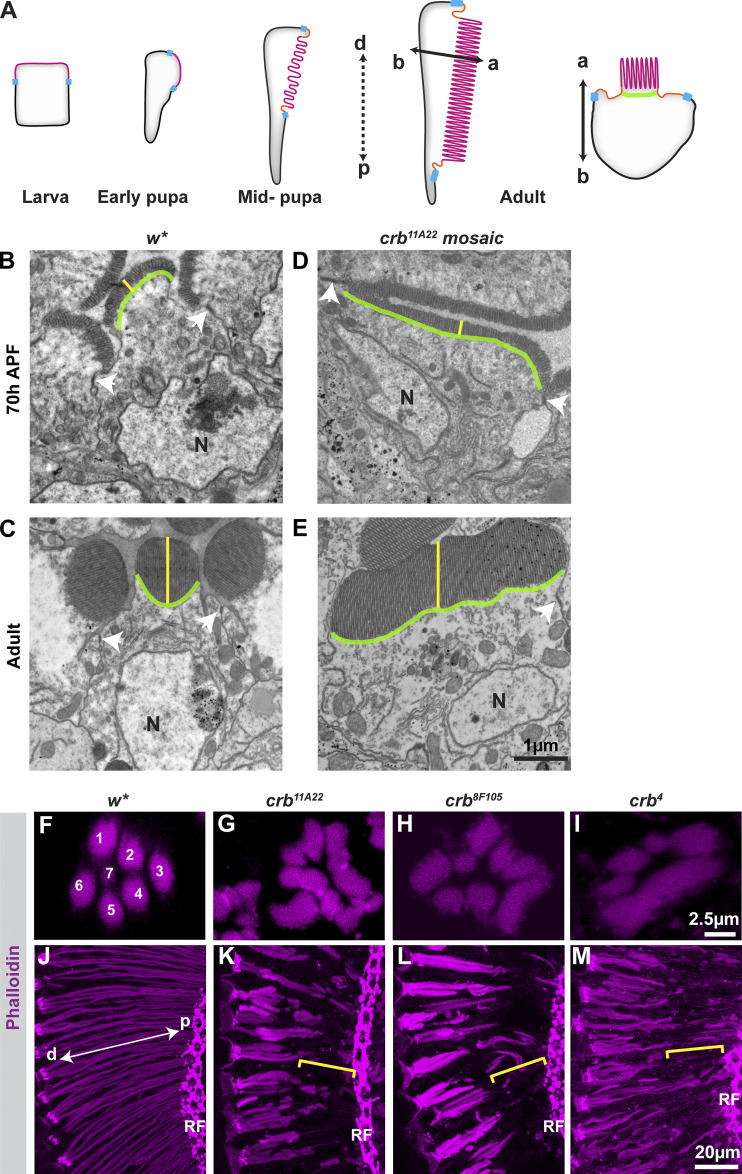
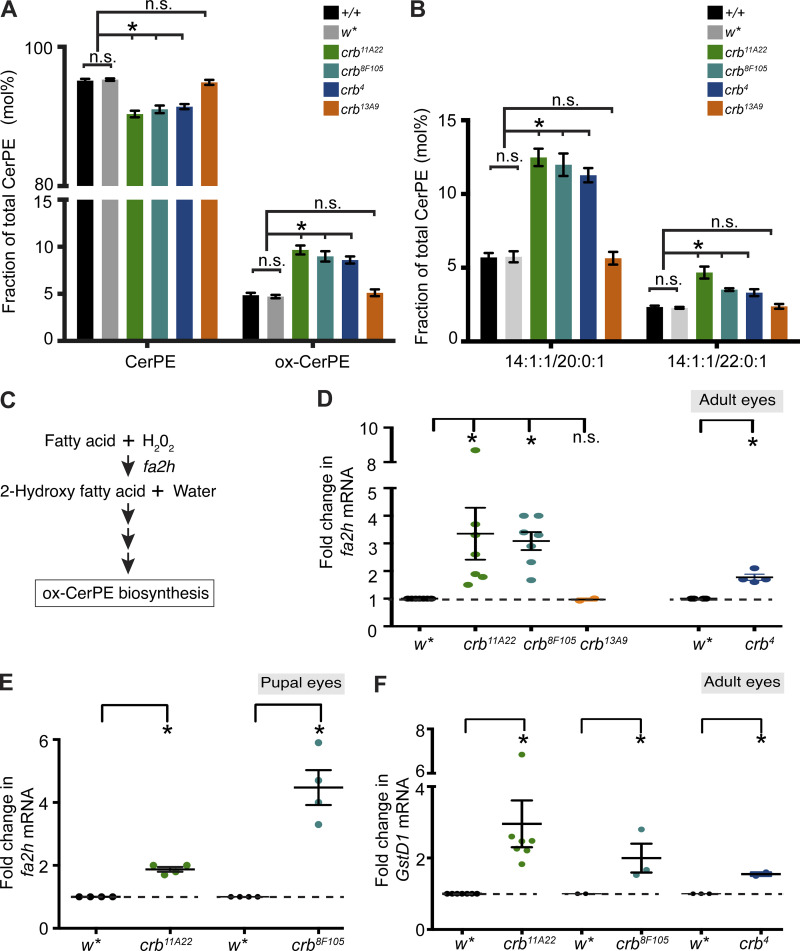
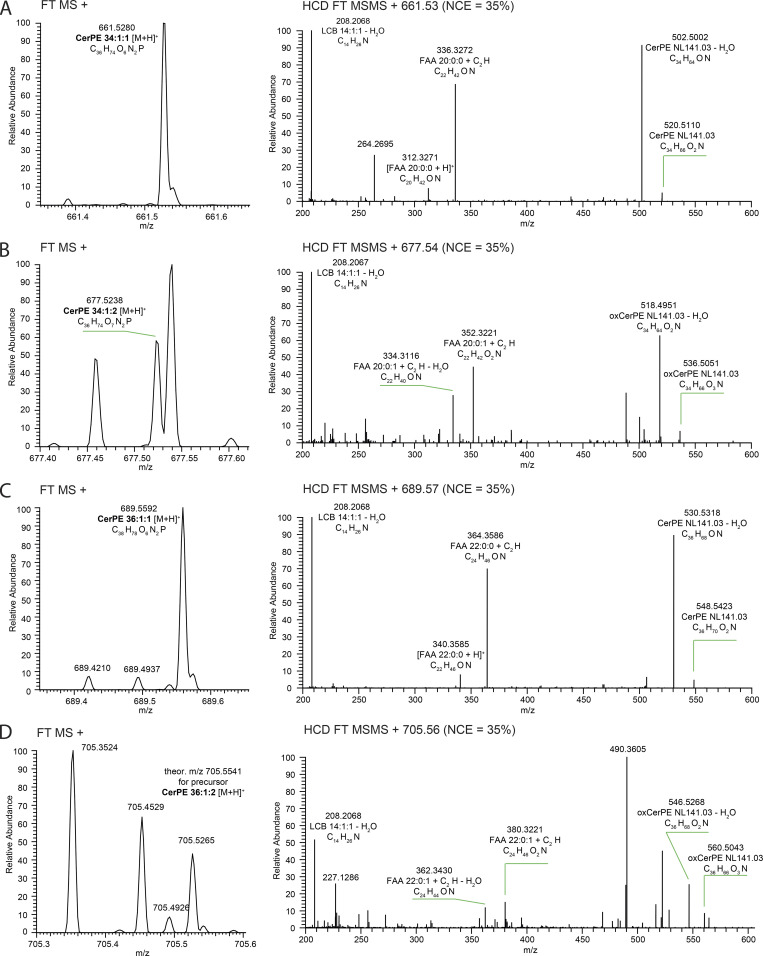
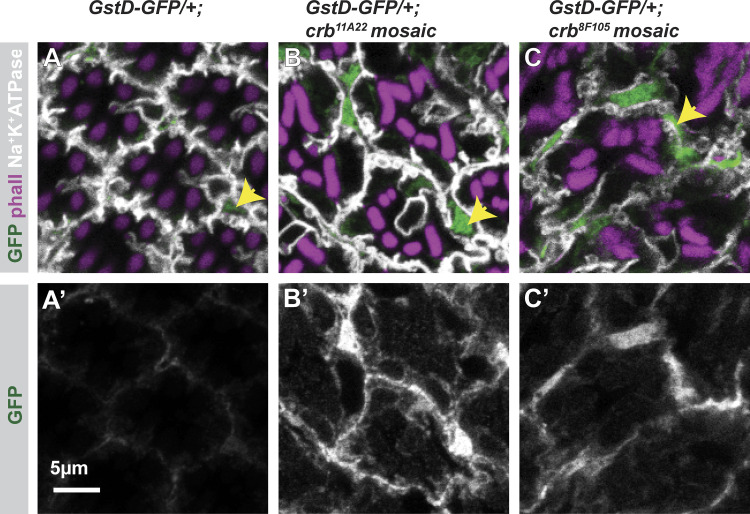
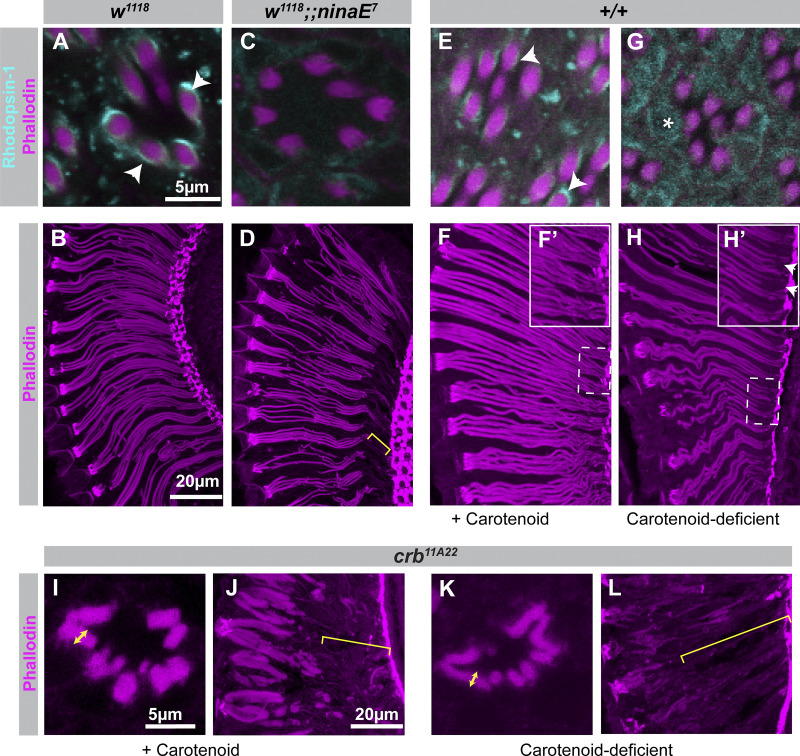
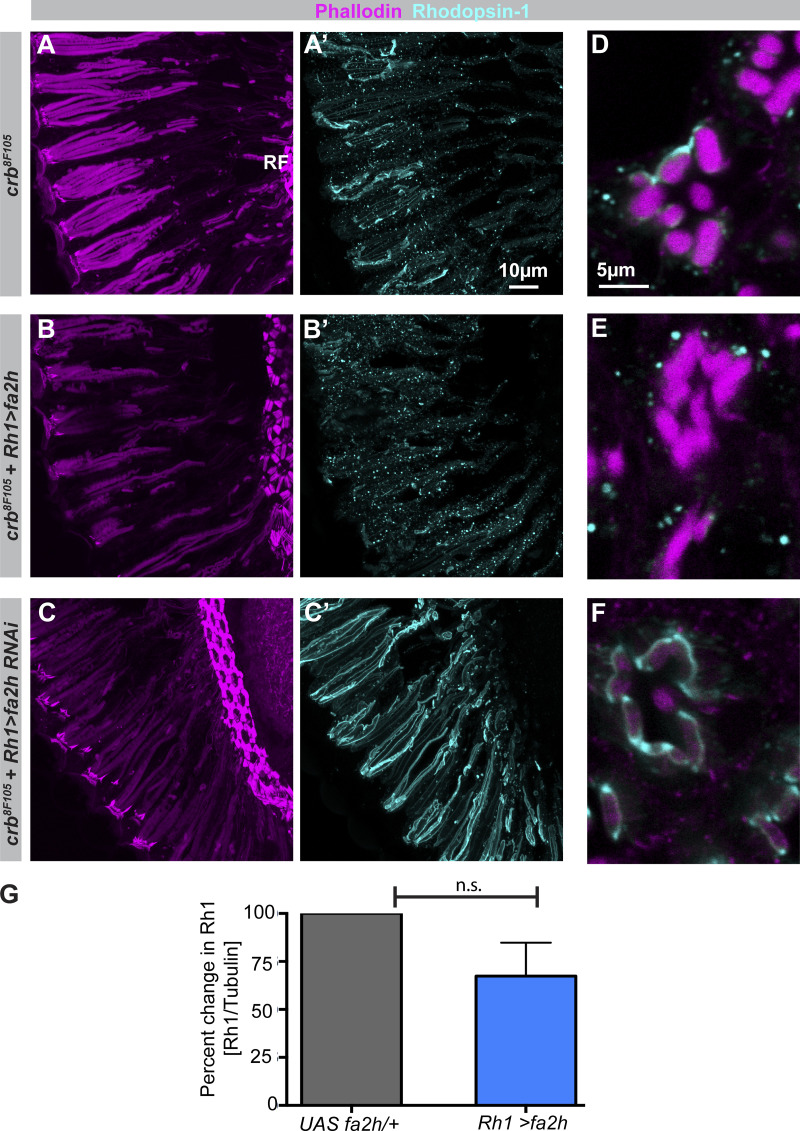
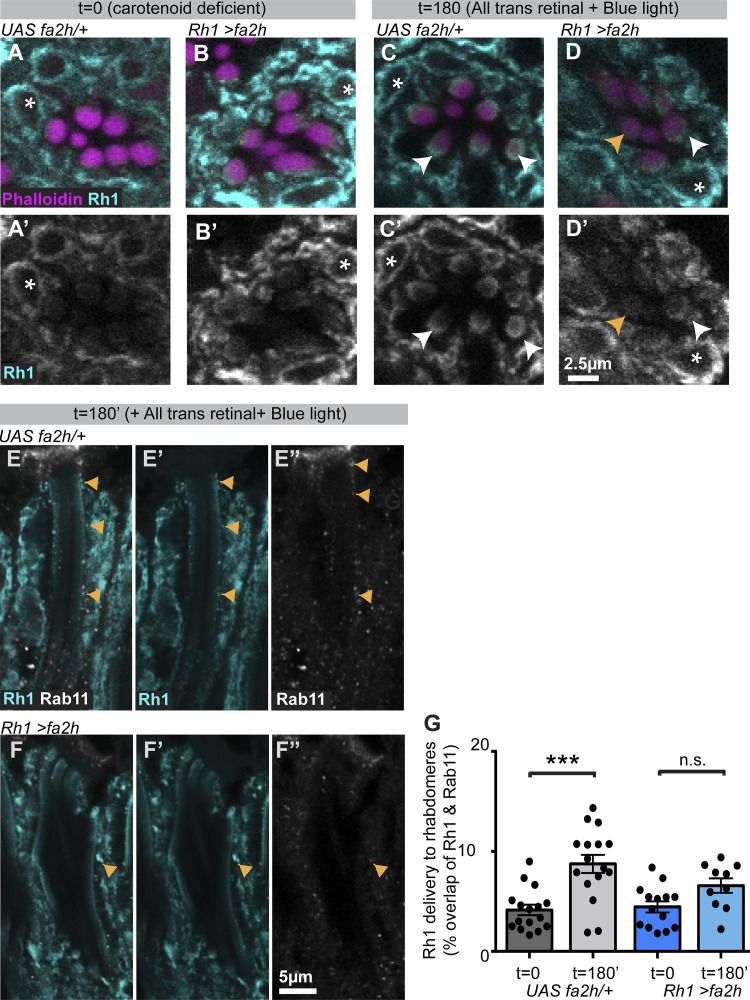
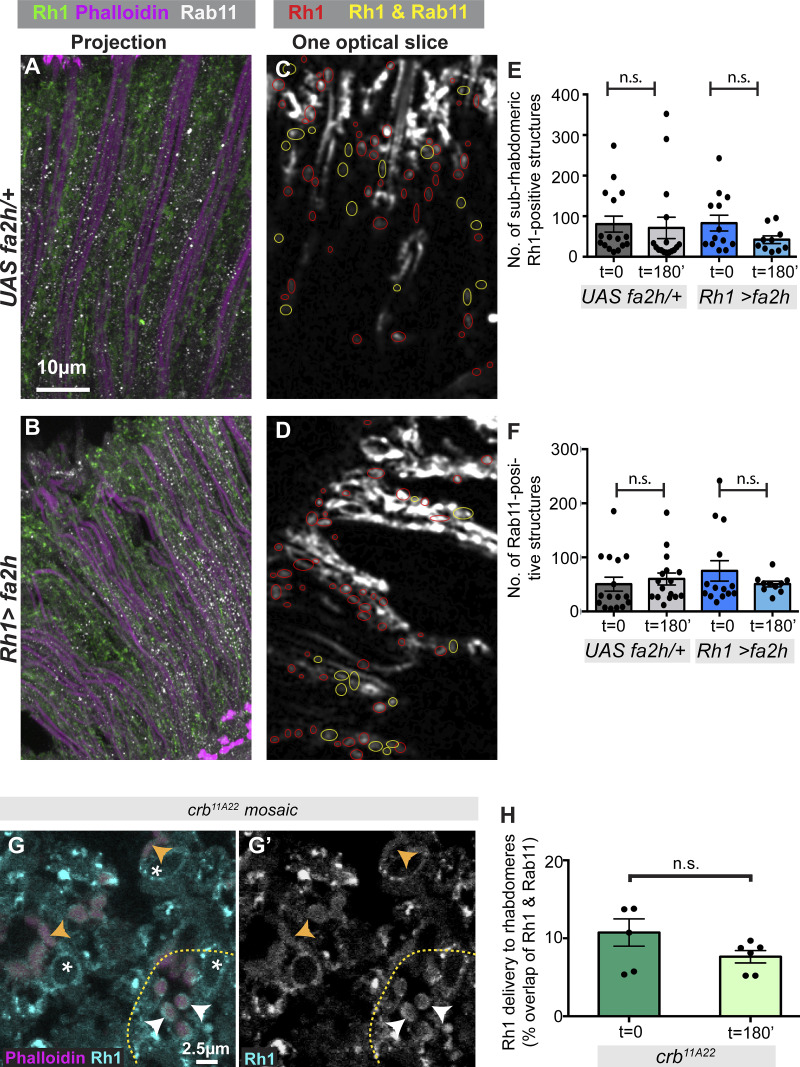
Apical domains of epithelial cells often undergo dramatic changes during morphogenesis to form specialized structures, such as microvilli. Here, we addressed the role of lipids during morphogenesis of the rhabdomere, the microvilli-based photosensitive organelle of Drosophila photoreceptor cells. Shotgun lipidomics analysis performed on mutant alleles of the polarity regulator crumbs, exhibiting varying rhabdomeric growth defects, revealed a correlation between increased abundance of hydroxylated sphingolipids and abnormal rhabdomeric growth. This could be attributed to an up-regulation of fatty acid hydroxylase transcription. Indeed, direct genetic perturbation of the hydroxylated sphingolipid metabolism modulated rhabdomere growth in a crumbs mutant background. One of the pathways targeted by sphingolipid metabolism turned out to be the secretory route of newly synthesized Rhodopsin, a major rhabdomeric protein. In particular, altered biosynthesis of hydroxylated sphingolipids impaired apical trafficking via Rab11, and thus apical membrane growth. The intersection of lipid metabolic pathways with apical domain growth provides a new facet to our understanding of apical growth during morphogenesis.
© 2020 Hebbar et al.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
