

Role of Glutathione in Cancer: From Mechanisms to Therapies
- PMID: 33050144
- PMCID: PMC7600400
- DOI: 10.3390/biom10101429
Role of Glutathione in Cancer: From Mechanisms to Therapies
Abstract
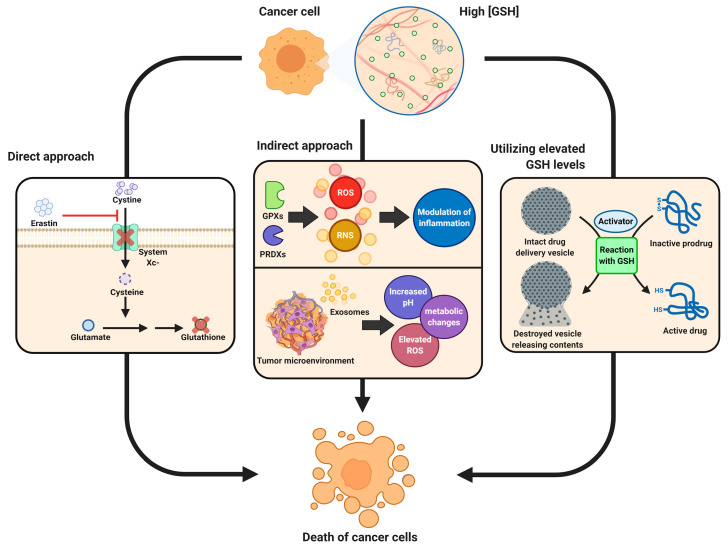
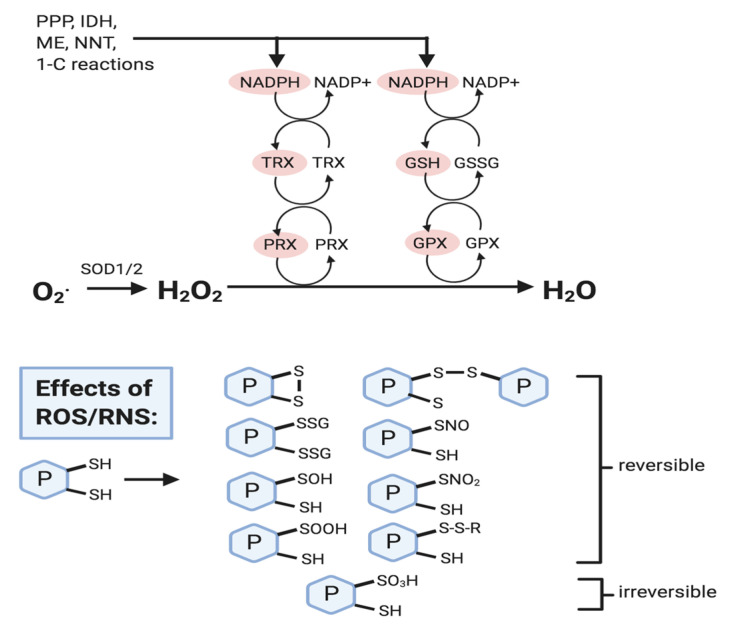
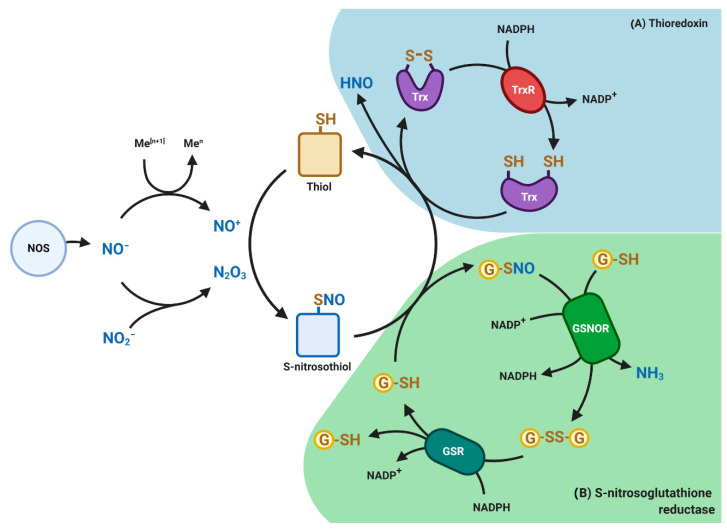
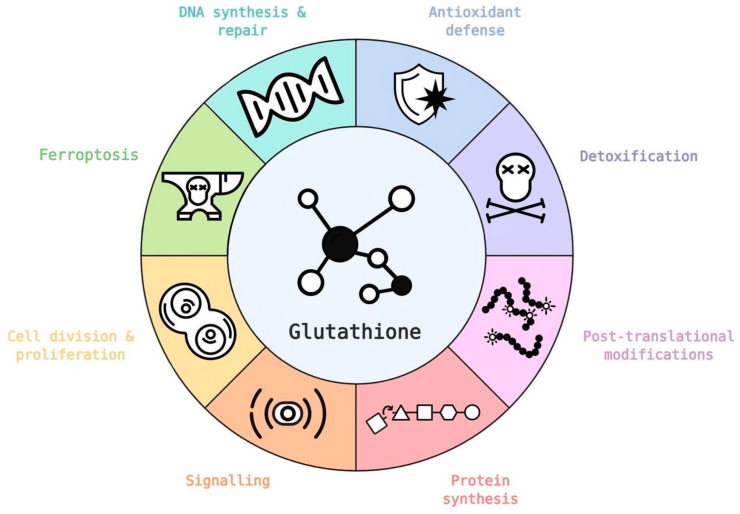
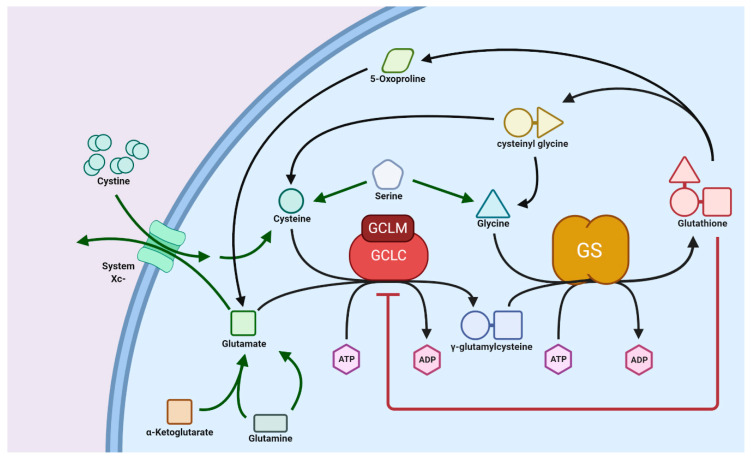
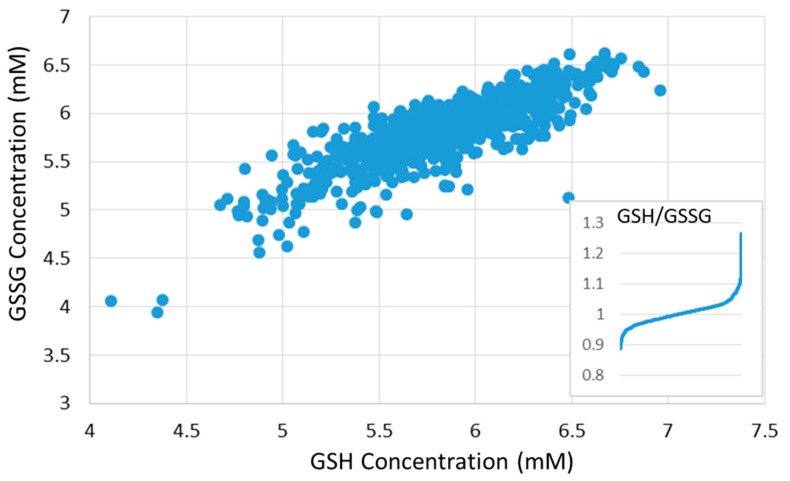
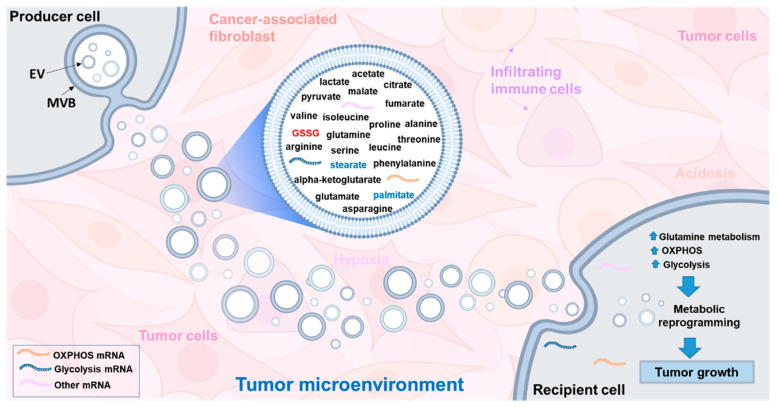
Glutathione (GSH) is the most abundant non-protein thiol present at millimolar concentrations in mammalian tissues. As an important intracellular antioxidant, it acts as a regulator of cellular redox state protecting cells from damage caused by lipid peroxides, reactive oxygen and nitrogen species, and xenobiotics. Recent studies have highlighted the importance of GSH in key signal transduction reactions as a controller of cell differentiation, proliferation, apoptosis, ferroptosis and immune function. Molecular changes in the GSH antioxidant system and disturbances in GSH homeostasis have been implicated in tumor initiation, progression, and treatment response. Hence, GSH has both protective and pathogenic roles. Although in healthy cells it is crucial for the removal and detoxification of carcinogens, elevated GSH levels in tumor cells are associated with tumor progression and increased resistance to chemotherapeutic drugs. Recently, several novel therapies have been developed to target the GSH antioxidant system in tumors as a means for increased response and decreased drug resistance. In this comprehensive review we explore mechanisms of GSH functionalities and different therapeutic approaches that either target GSH directly, indirectly or use GSH-based prodrugs. Consideration is also given to the computational methods used to describe GSH related processes for in silico testing of treatment effects.
Keywords: S-nitrosation; ferroptosis; glutathione; metabolism modeling; nitrosoglutathione; reactive oxygen species; tumor metabolism; tumor therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
