

Molecular and Cellular Pathogenesis of Ellis-van Creveld Syndrome: Lessons from Targeted and Natural Mutations in Animal Models
- PMID: 33050204
- PMCID: PMC7711556
- DOI: 10.3390/jdb8040025
Molecular and Cellular Pathogenesis of Ellis-van Creveld Syndrome: Lessons from Targeted and Natural Mutations in Animal Models
Abstract
Ellis-van Creveld syndrome (EVC; MIM ID #225500) is a rare congenital disease with an occurrence of 1 in 60,000. It is characterized by remarkable skeletal dysplasia, such as short limbs, ribs and polydactyly, and orofacial anomalies. With two of three patients first noted as being offspring of consanguineous marriage, this autosomal recessive disease results from mutations in one of two causative genes: EVC or EVC2/LIMBIN. The recent identification and manipulation of genetic homologs in animals has deepened our understanding beyond human case studies and provided critical insight into disease pathogenesis. This review highlights the utility of animal-based studies of EVC by summarizing: (1) molecular biology of EVC and EVC2/LIMBIN, (2) human disease signs, (3) dysplastic limb development, (4) craniofacial anomalies, (5) tooth anomalies, (6) tracheal cartilage abnormalities, and (7) EVC-like disorders in non-human species.
Keywords: EVC2; Ellis-van Creveld syndrome; LIMBIN; ciliopathy; craniofacial.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

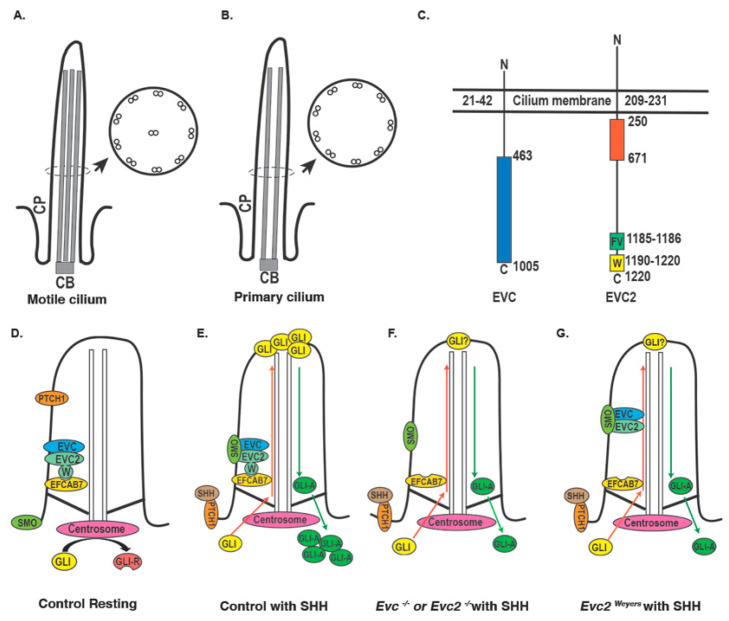
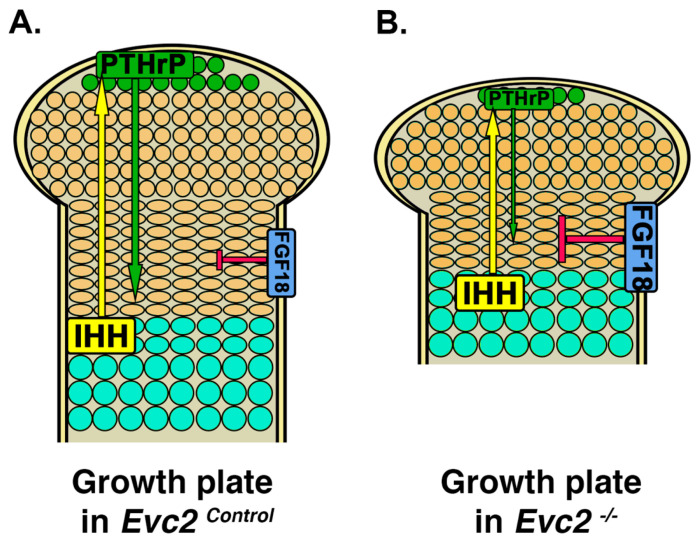
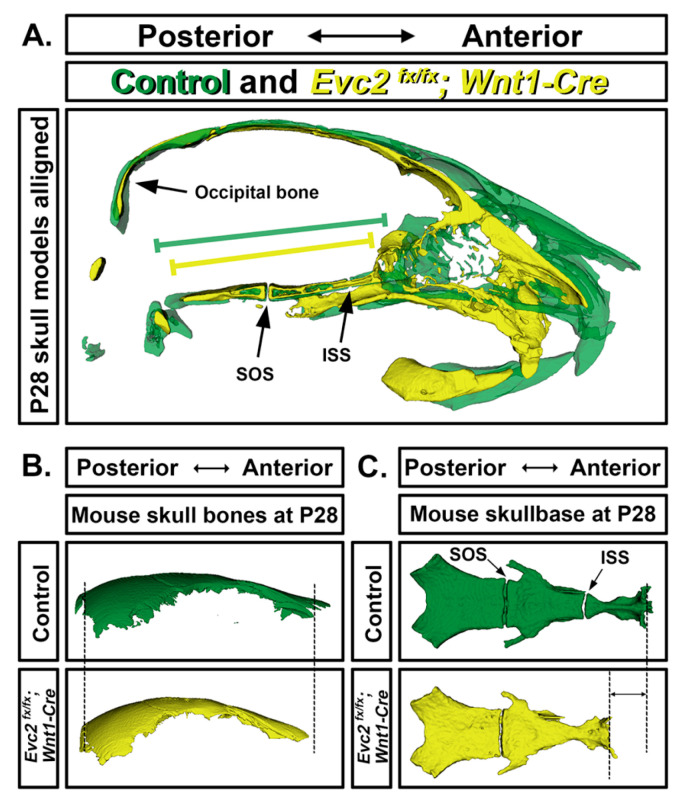
Similar articles
-
Elevated Fibroblast Growth Factor Signaling Is Critical for the Pathogenesis of the Dwarfism in Evc2/Limbin Mutant Mice.PLoS Genet. 2016 Dec 27;12(12):e1006510. doi: 10.1371/journal.pgen.1006510. eCollection 2016 Dec. PLoS Genet. 2016. PMID: 28027321 Free PMC article.
-
Novel and recurrent EVC and EVC2 mutations in Ellis-van Creveld syndrome and Weyers acrofacial dyostosis.Eur J Med Genet. 2013 Feb;56(2):80-7. doi: 10.1016/j.ejmg.2012.11.005. Epub 2012 Dec 7. Eur J Med Genet. 2013. PMID: 23220543
-
Role of primary cilia and Hedgehog signaling in craniofacial features of Ellis-van Creveld syndrome.Am J Med Genet C Semin Med Genet. 2022 Mar;190(1):36-46. doi: 10.1002/ajmg.c.31969. Epub 2022 Apr 8. Am J Med Genet C Semin Med Genet. 2022. PMID: 35393766 Review.
-
Novel homozygous mutations in the EVC and EVC2 genes in two consanguineous families segregating autosomal recessive Ellis-van Creveld syndrome.Clin Dysmorphol. 2016 Jan;25(1):1-6. doi: 10.1097/MCD.0000000000000104. Clin Dysmorphol. 2016. PMID: 26580685
-
Ellis-van Creveld syndrome and Weyers acrodental dysostosis are caused by cilia-mediated diminished response to hedgehog ligands.Am J Med Genet C Semin Med Genet. 2009 Nov 15;151C(4):341-51. doi: 10.1002/ajmg.c.30226. Am J Med Genet C Semin Med Genet. 2009. PMID: 19876929 Review.
Cited by
-
Novel Candidate Genes for Non-Syndromic Tooth Agenesis Identified Using Targeted Next-Generation Sequencing.J Clin Med. 2022 Oct 15;11(20):6089. doi: 10.3390/jcm11206089. J Clin Med. 2022. PMID: 36294409 Free PMC article.
-
The Posterior Part Influences the Anterior Part of the Mouse Cranial Base Development.JBMR Plus. 2021 Dec 24;6(2):e10589. doi: 10.1002/jbm4.10589. eCollection 2022 Feb. JBMR Plus. 2021. PMID: 35229066 Free PMC article.
-
Establishing an objective clinical spectrum, genotype-phenotype correlations, and CRMP1 as a modifier in the Ellis-van Creveld syndrome: The first systematic review of EVC- and EVC2-associated conditions.Genet Med Open. 2023 Mar 13;1(1):100781. doi: 10.1016/j.gimo.2023.100781. eCollection 2023. Genet Med Open. 2023. PMID: 39669252 Free PMC article. Review.
-
Skeletal ciliopathy: pathogenesis and related signaling pathways.Mol Cell Biochem. 2024 Apr;479(4):811-823. doi: 10.1007/s11010-023-04765-5. Epub 2023 May 15. Mol Cell Biochem. 2024. PMID: 37188988 Review.
-
Identification of Compound Heterozygous EVC2 Gene Variants in Two Mexican Families with Ellis-van Creveld Syndrome.Genes (Basel). 2023 Apr 9;14(4):887. doi: 10.3390/genes14040887. Genes (Basel). 2023. PMID: 37107645 Free PMC article.
References
-
- McKusick V.A. Dwarfism in the Amish I. The Ellis-van Creveld Syndrome. Bull. Johns Hopkins Hosp. 1964;115:306–336. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
