

p31comet promotes homologous recombination by inactivating REV7 through the TRIP13 ATPase
- PMID: 33051298
- PMCID: PMC7604461
- DOI: 10.1073/pnas.2008830117
p31comet promotes homologous recombination by inactivating REV7 through the TRIP13 ATPase
Abstract
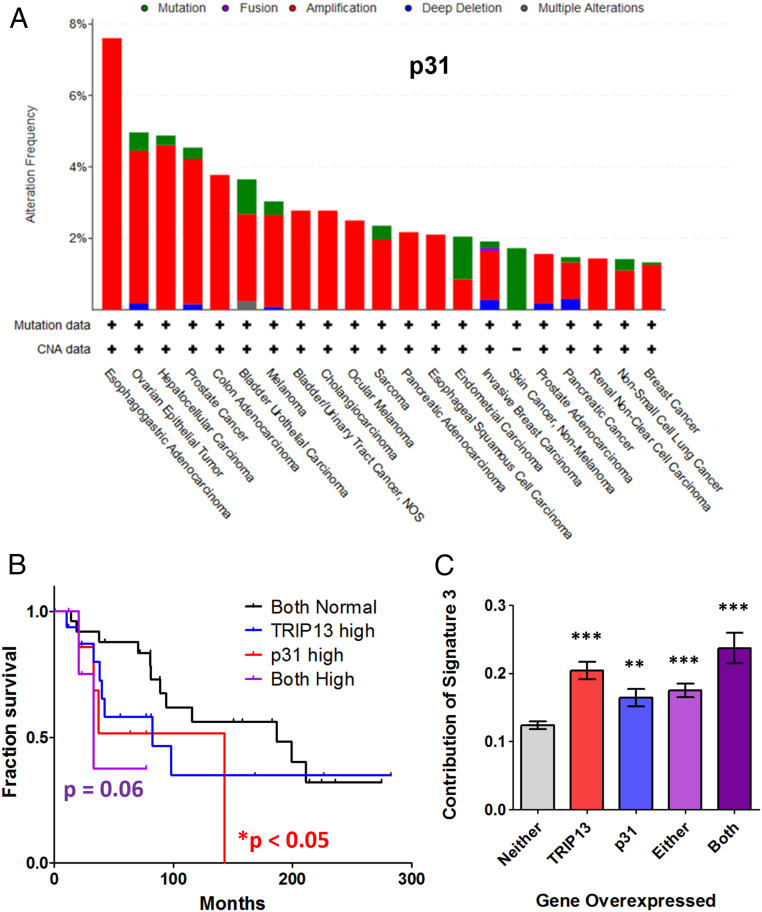
The repair of DNA double strand breaks (DSBs) that arise from external mutagenic agents and routine cellular processes is essential for life. DSBs are repaired by two major pathways, homologous recombination (HR) and classical nonhomologous end joining (C-NHEJ). DSB repair pathway choice is largely dictated at the step of 5'-3' DNA end resection, which is promoted during S phase, in part by BRCA1. Opposing end resection is the 53BP1 protein, which recruits the ssDNA-binding REV7-Shieldin complex to favor C-NHEJ repair. We recently identified TRIP13 as a proresection factor that remodels REV7, causing its dissociation from the Shieldin subunit SHLD3. Here, we identify p31comet, a negative regulator of MAD2 and the spindle assembly checkpoint, as an important mediator of the TRIP13-REV7 interaction. p31comet binds to the REV7-Shieldin complex in cells, promotes REV7 inactivation, and causes PARP inhibitor resistance. p31comet also participates in the extraction of REV7 from the chromatin. Furthermore, p31comet can counteract REV7 function in translesion synthesis (TLS) by releasing it from REV3 in the Pol ζ complex. Finally, p31comet, like TRIP13, is overexpressed in many cancers and this correlates with poor prognosis. Thus, we reveal a key player in the regulation of HR and TLS with significant clinical implications.
Keywords: Fanconi anemia; PARP inhibitor; REV7; homologous recombination; translesion synthesis.
Copyright © 2020 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: A.D.D. is a consultant/advisory board member for Lilly Oncology, Merck-EMD Serono, Intellia Therapeutics, Sierra Oncology, Cyteir Therapeutics, Third Rock Ventures, AstraZeneca, Ideaya Inc., Cedilla Therapeutics Inc., a stockholder in Ideaya Inc., Cedilla Therapeutics Inc., and Cyteir, and reports receiving commercial research grants from Lilly Oncology and Merck-EMD Serono.
Figures
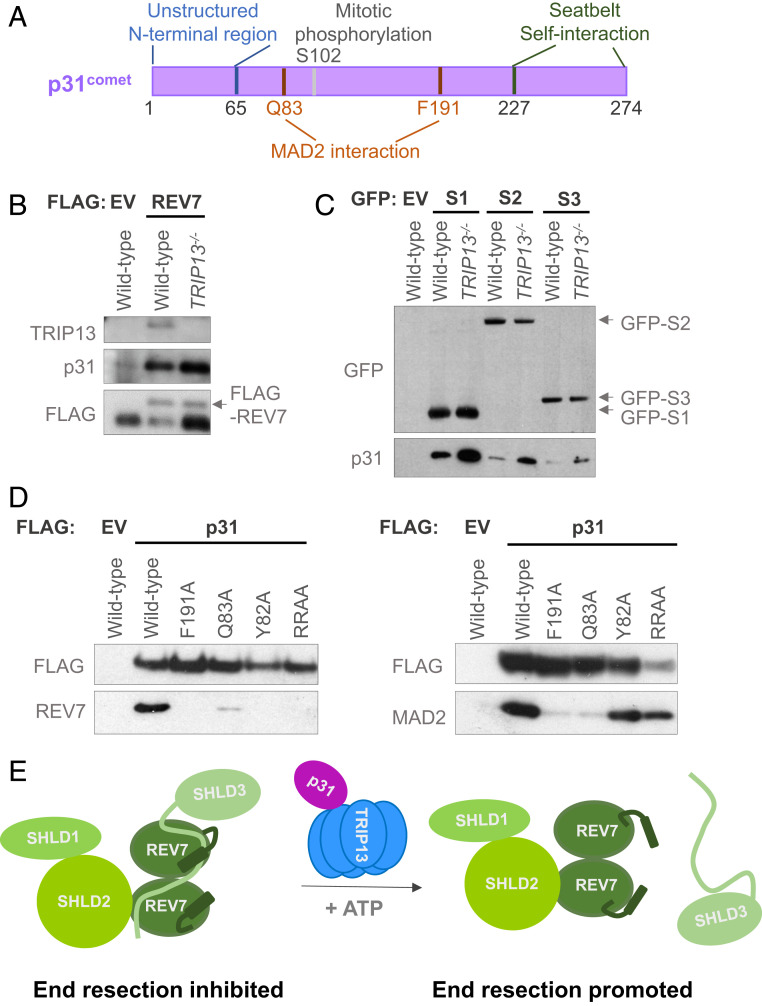
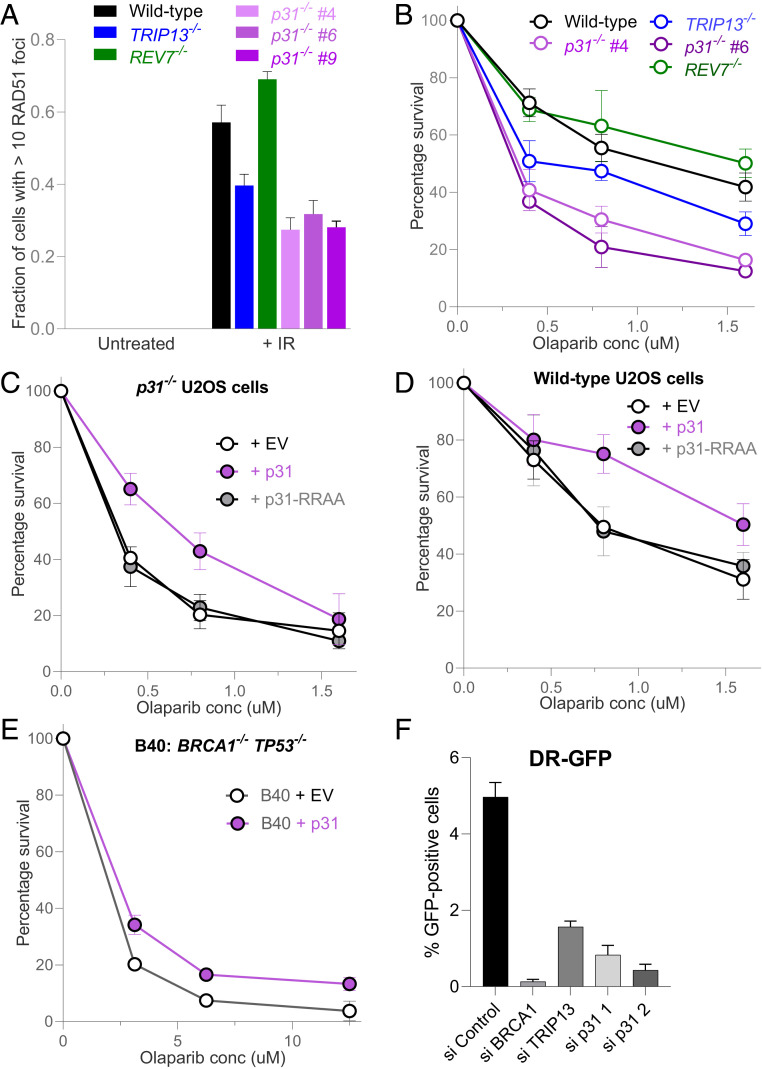
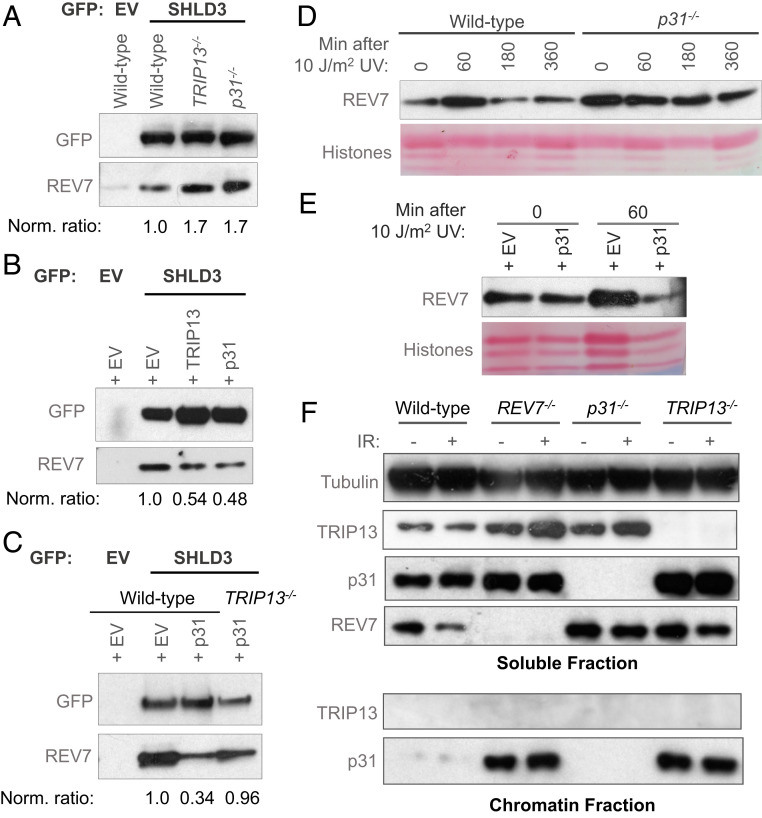
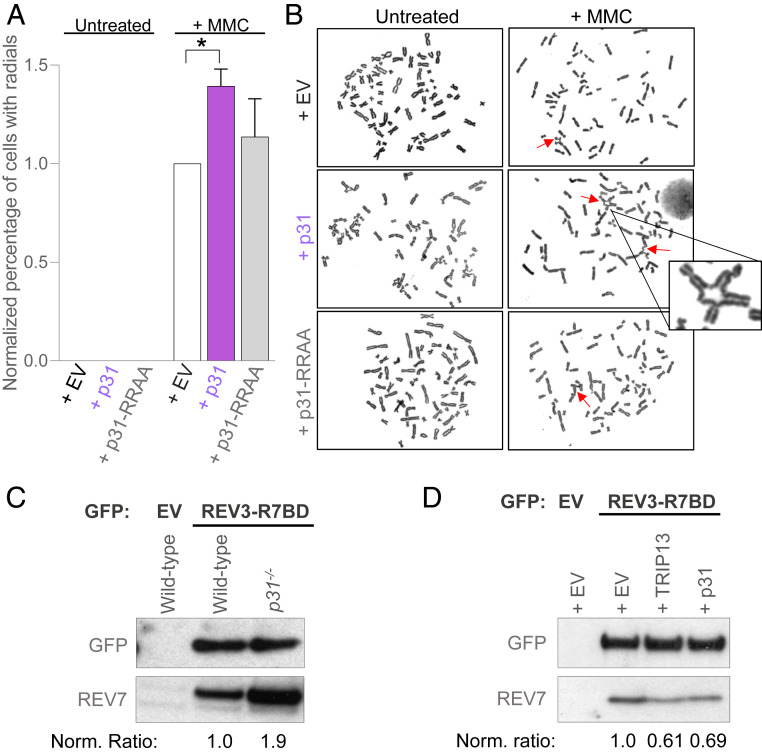
Comment in
-
p31comet and TRIP13 recycle Rev7 to regulate DNA repair.Proc Natl Acad Sci U S A. 2020 Nov 10;117(45):27761-27763. doi: 10.1073/pnas.2020103117. Epub 2020 Oct 29. Proc Natl Acad Sci U S A. 2020. PMID: 33122436 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
