

Review
doi: 10.1152/physiol.00015.2020.
Multi-Level Regulation of Opioid-Induced Respiratory Depression
Affiliations
- PMID: 33052772
- PMCID: PMC7864237
- DOI: 10.1152/physiol.00015.2020
Item in Clipboard
Review
Multi-Level Regulation of Opioid-Induced Respiratory Depression
Physiology (Bethesda).
.
Abstract
Opioids depress minute ventilation primarily by reducing respiratory rate. This results from direct effects on the preBötzinger Complex as well as from depression of the Parabrachial/Kölliker-Fuse Complex, which provides excitatory drive to preBötzinger Complex neurons mediating respiratory phase-switch. Opioids also depress awake drive from the forebrain and chemodrive.
Keywords: Parabrachial Nucleus/Kölliker-Fuse Complex; awake drive; chemodrive; opioids; preBötzinger Complex.
Figures
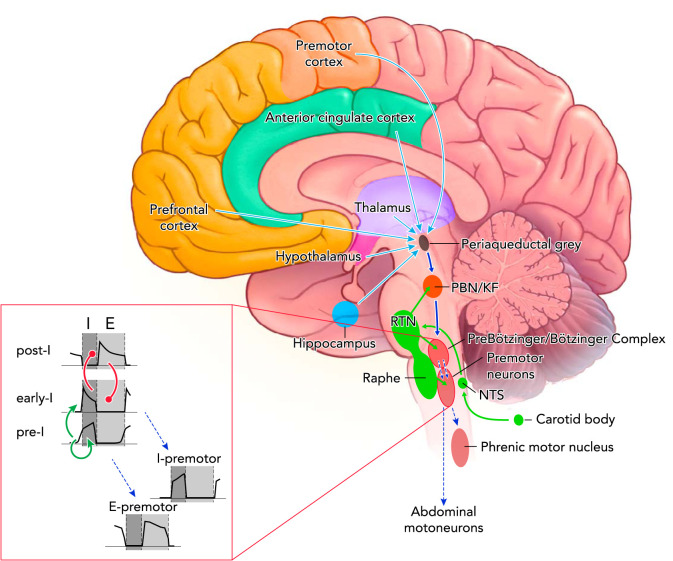
Excitatory connections within the respiratory control center and opioid effects Tonic chemodrive (green solid arrows) is the main excitatory drive to the medullary rhythm generator. A large component of chemodrive is routed through the Parabrachial Nucleus/Kölliker-Fuse Complex to phase-switching neurons in the preBötzinger Complex and determines respiratory rate (dark blue solid arrow). Phasic inputs from the preBötzinger Complex activate inspiratory (I) and expiratory (E) premotor and motoneurons (blue dotted arrows). Respiratory motor output (tidal volume) depends on direct projections from the retrotrapezoid nucleus to preBötzinger Complex neurons, premotor neurons, and motoneurons. The cortico-limbic system contributes tonic drive to the medullary rhythm generator (light blue solid arrows). Not shown: direct projections from the motor cortex to phrenic motoneurons as a pathway to override automatic rhythm; projections from the hypothalamus and cerebellum to the medullary raphe, which may contribute to the state-dependency of respiratory activity. Opioid effects: Parabrachial Nucleus/Kölliker-Fuse Complex activity was depressed in all studies and at all opioid concentrations. Opioid-induced depression was also shown for the preBötzinger Complex, the nucleus of the solitary tract, and the medullary raphe. Opioid-induced sedation suggests depression of forebrain inputs. Premotor neurons are only directly depressed at very high opioid concentrations. Inset: neuronal subtypes constituting the core of respiratory rhythm generation. Through mutual excitation, network activity of pre-inspiratory neurons (pre-I) in the preBötzinger Complex results in activation of early inspiratory (early-I) neurons (green arrows). Activity of these neurons is terminated through inhibition by post-inspiratory neurons (post-I), which themselves are inhibited by early-I neurons during the inspiratory phase (red circles). Phasic excitation is relayed to inspiratory (I) and expiratory (E) premotor neurons.
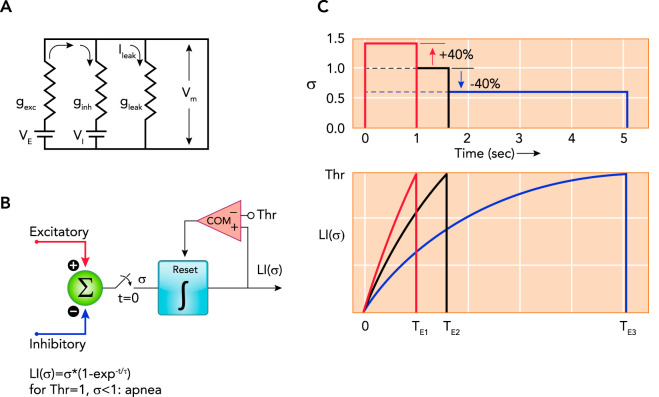
Model of the components determining the membrane potential and illustrating the effects of changes in excitatory and/or inhibitory inputs A: model of the components determining the membrane potential (Vm) of a preBötzinger Complex phase-switching neuron: , where gexc is excitatory conductance, VE is equilibrium potential for excitatory currents, ginh is inhibitory conductance, VI is equilibrium potential for inhibitory currents, and gleak is leak conductance, e.g., GIRK channels (see text for details). B and C: functional timer model to illustrate the effects of changes in excitatory and/or inhibitory inputs on the neuronal membrane trajectory (see Refs. , , –173). B: the sum of all excitatory and inhibitory inputs (σ) is gated to a leaky integrator (LI) at time, t = 0 s. The magnitude of σ determines the rate of the exponential rise of Vm to a threshold (Thr). Crossing the threshold resets the leaky integrator via a comparator (COM). The time to crossing the threshold determines the phase duration. C: graphic illustration of the timing operation using the example of rhythmogenic pre-inspiratory neurons. Neuronal discharge in pre-inspiratory neurons begins when Thr is reached and results in inspiratory on-switch. Inspiratory on-switch terminates the expiratory phase, i.e., the time to Thr for pre-inspiratory neurons determines expiratory duration (TE). Due to the nonlinear nature of this mechanism, increases and decreases in σ of the same magnitude cause strikingly different changes in phase duration. Shown are three examples for σ (upper) and the corresponding leaky integrator outputs LI (σ) (lower). Setting σ = 1.0 as a baseline reference results in a duration of TE2 = 1.6 s (black lines). Increasing σ by 40% results in TE1 = 1 s (red), whereas decreasing σ by 40% results in TE3 = 5 s (blue). Physiological examples for an increase in σ could be an increase in neuronal activity in the parabrachial nucleus (PBN)/Kölliker-Fuse nucleus (137) or an increase in inhibitory activity during the preceding inspiratory phase, which shortens inspiratory duration and presumably shortens the post-inspiratory refractory period (3). The latter could be due to vagal pulmonary stretch receptor input during lung inflation or increased activity of preBötzinger Complex inhibitory neurons during the inspiratory phase (3). Decreases in σ could be due to a reduction in PBN activity via inhibition of PBN neurons by opioids, by increases in pulmonary stretch receptor activity during the expiratory phase (173), or by increases in preBötzinger Complex GABAergic/glycinergic neuronal activity during the expiratory phase (3).
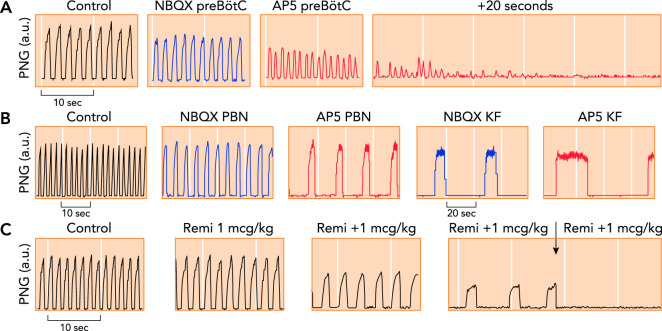
Phrenic neurogram tracings obtained in adult decerebrate rabbits in vivo with Pco 2 constant A: antagonism of glutamate receptor function in the preBötzinger Complex (preBötC) through local microinjection of the AMPA receptor antagonist NBQX (blue) and the NMDA receptor antagonist AP5 (red) caused tachypnea with decreased peak phrenic amplitude, followed by apnea (25). B: antagonism of glutamate receptor function in the parabrachial nucleus (PBN) and Kölliker-Fuse nucleus (KF) through local microinjection of NBQX (blue) and AP5 (red) caused severe bradypnea, whereas peak phrenic amplitude was only little decreased. Complete apnea was never observed (116). Please note the different time scales. C: repeated intravenous boluses of the mu-opioid agonist remifentanil (Remi; 1 mcg · kg–1
bolus–1) caused increasing bradypnea due to prolongation of inspiratory and expiratory phase duration. The fourth bolus resulted in apnea (arrow). Remifentanil also substantially depressed peak phrenic activity (123), suggesting an effect on both the PBN/KF and the preBötC.
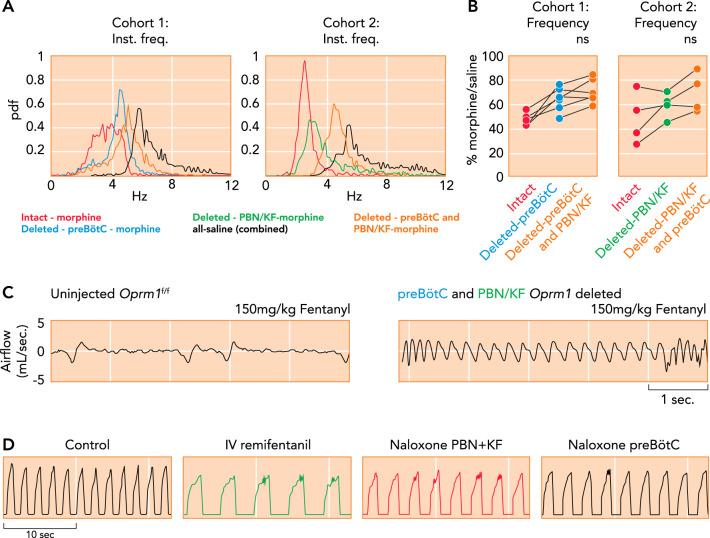
Critical importance of the Parabrachial Nucleus/Kölliker-Fuse complex and preBötzinger Complex for opioid-induced respiratory depression A–C: μ-opioid receptors (Oprm1) were selectively deleted in Oprm1
f/f mice through Cre-virus injection into the preBötC and PBN/KF in this (cohort 1) or reverse order (cohort 2) with 4–5 wk between injections and plethysmography recordings (data from A–C from Ref , and used with permission from eLife). A: probability density function plot of the respiratory rate for a representative animal from cohorts 1 and 2 after intraperitoneal morphine (20 mg/kg) or saline before and after Oprm1 deletion in the PBN/KF and preBötC. B: pooled data showed that morphine-induced respiratory rate depression was significantly attenuated after Oprm1 deletion in the preBötC. Oprm1 deletion in both areas reduced morphine-induced respiratory rate depression from ~50% to ~30%. C: plethysmography recordings in mice show that Oprm1 deletion in the preBötC + PBN/KF prevented respiratory rate depression from a very high intraperitoneal fentanyl dose (150 mg/kg; right) that usually caused lethal apnea before Oprm1 deletion (left). D: consistent with the murine studies, phrenic neurogram tracings obtained in an adult decerebrate rabbit in vivo show that sequential microinjections of the opioid antagonist naloxone into the bilateral PBN + KF and bilateral preBötC reversed the respiratory rate depression from intravenous remifentanil infusion (see Ref. 123).
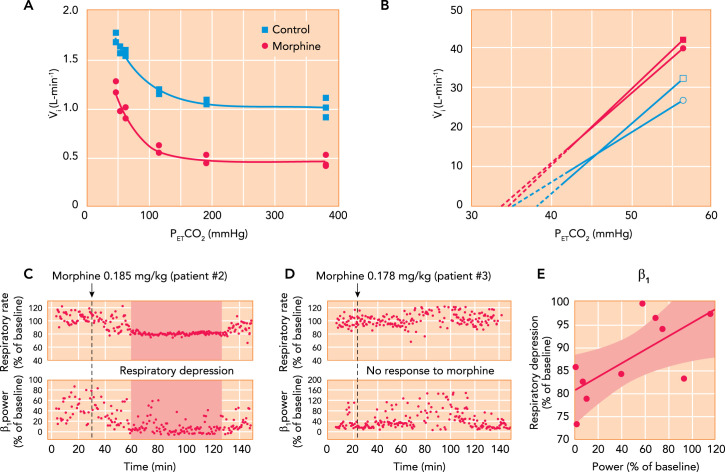
Contributions of chemodrive and “awake drive” to minute ventilation and opioid effects A: hypoxic ventilatory response curves in a chloralose-urethane anesthetized cat during control (solid square) and after administration of 0.15 mg/kg IV morphine. Morphine decreased minute ventilation during hyperoxia but did not change the increase in minute ventilation with hypoxia (see Ref. 12). B: mean ventilatory response to increasing inspiratory carbon dioxide concentrations obtained in 12 male (square) and 12 female (circle) human volunteers during control (filled) and after 0.1 mg/kg IV morphine, followed by 0.03 mg · kg–1 · h–1 (open). The continuous lines are the linear regression lines through the data points, and broken lines are extrapolated to the apneic threshold. In men and women, morphine decreased minute ventilation differently: in men by increasing the apneic threshold but in women by decreasing carbon dioxide sensitivity (see Ref. 30). C–E: association between respiratory depression from analgesic doses of morphine and loss of “awake drive” per electroencephalogram in pediatric patients. C: after 0.185 mg/kg morphine, patient 2 presented substantial respiratory rate depression associated with a decrease in β1 power. D: in patient 3, a similar dose (0.178 mg/kg) did not reduce β1 power or cause notable respiratory depression. E: in 10 patients, the severity of respiratory rate depression correlated with the intensity of the reduction in β1 power (R = 0.715, P = 0.02) (105).
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical