

An isoform-specific pivot modulates the electron transfer between the flavin mononucleotide and heme centers in inducible nitric oxide synthase
- PMID: 33057871
- PMCID: PMC7669679
- DOI: 10.1007/s00775-020-01824-w
An isoform-specific pivot modulates the electron transfer between the flavin mononucleotide and heme centers in inducible nitric oxide synthase
Abstract
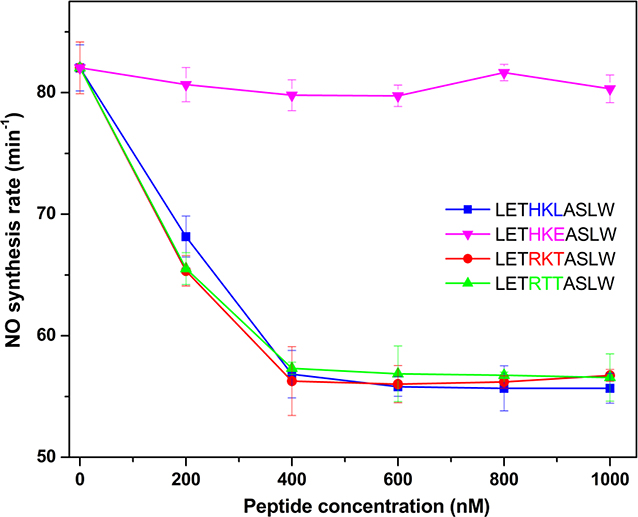
Intraprotein interdomain electron transfer (IET) between the flavin mononucleotide (FMN) and heme centers is an obligatory step in nitric oxide synthase (NOS) enzymes. An isoform-specific pivotal region near Leu406 in the heme domain of human inducible NOS (iNOS) was proposed to mediate the FMN-heme domain-domain alignment (J Inorg Biochem 153:186-196, 2015). The FMN-heme IET rate is a measure of the interdomain FMN/heme complex formation. In this work, the FMN-heme IET kinetics in the wild type (wt) human iNOS oxygenase/FMN (oxyFMN) construct were directly measured by laser flash photolysis with added synthetic peptide related to the pivotal region, in comparison with the wt construct alone. The IET rates were decreased by the iNOS HKL peptide in a dose-saturable fashion, and the inhibitory effect was abolished by a single L406 → E mutation in the peptide. A similar trend in change of the NO synthesis activity of wt iNOS holoenzyme by the peptides was observed. These data, along with the kinetics and modeling results for the L406T and L406F mutant oxyFMN proteins, indicated that the Leu406 residue modulates the FMN-heme IET through hydrophobic interactions. Moreover, the IET rates were analyzed for the wt iNOS oxyFMN protein in the presence of nNOS or eNOS-derived peptide related to the equivalent pivotal heme domain site. These results together indicate that the isoform-specific pivotal region at the heme domain specifically interacts with the conserved FMN domain surface, to facilitate proper interdomain docking for the FMN-heme IET in NOS.
Keywords: Electron transfer; Kinetics; Laser flash photolysis; Nitric oxide synthase.
Conflict of interest statement
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
