

A MicroRNA Linking Human Positive Selection and Metabolic Disorders
- PMID: 33058756
- PMCID: PMC8092355
- DOI: 10.1016/j.cell.2020.09.017
A MicroRNA Linking Human Positive Selection and Metabolic Disorders
Abstract
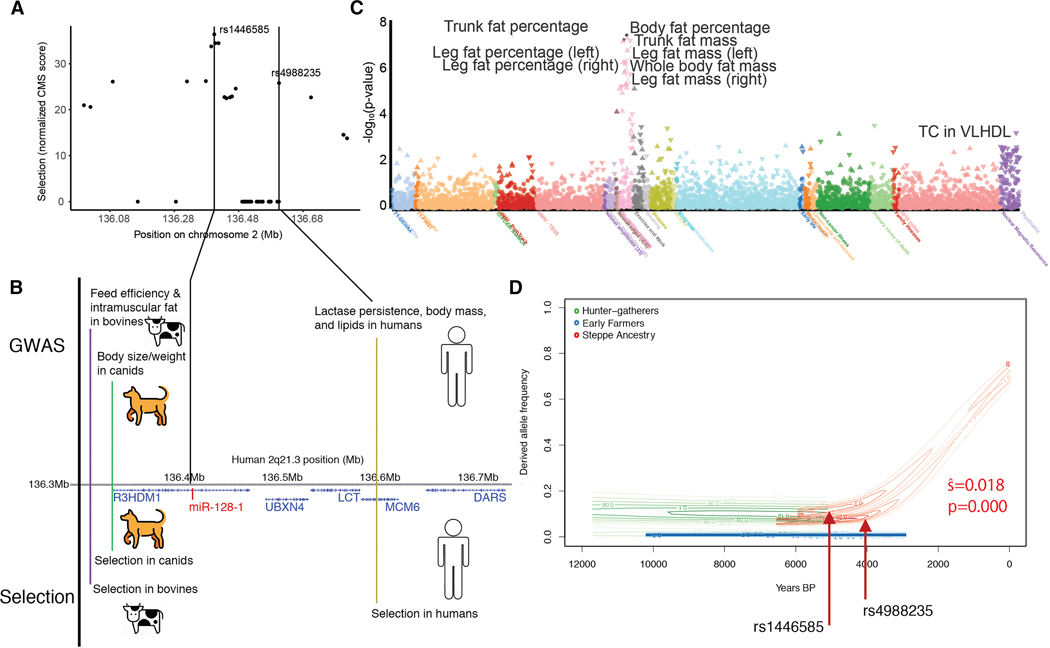
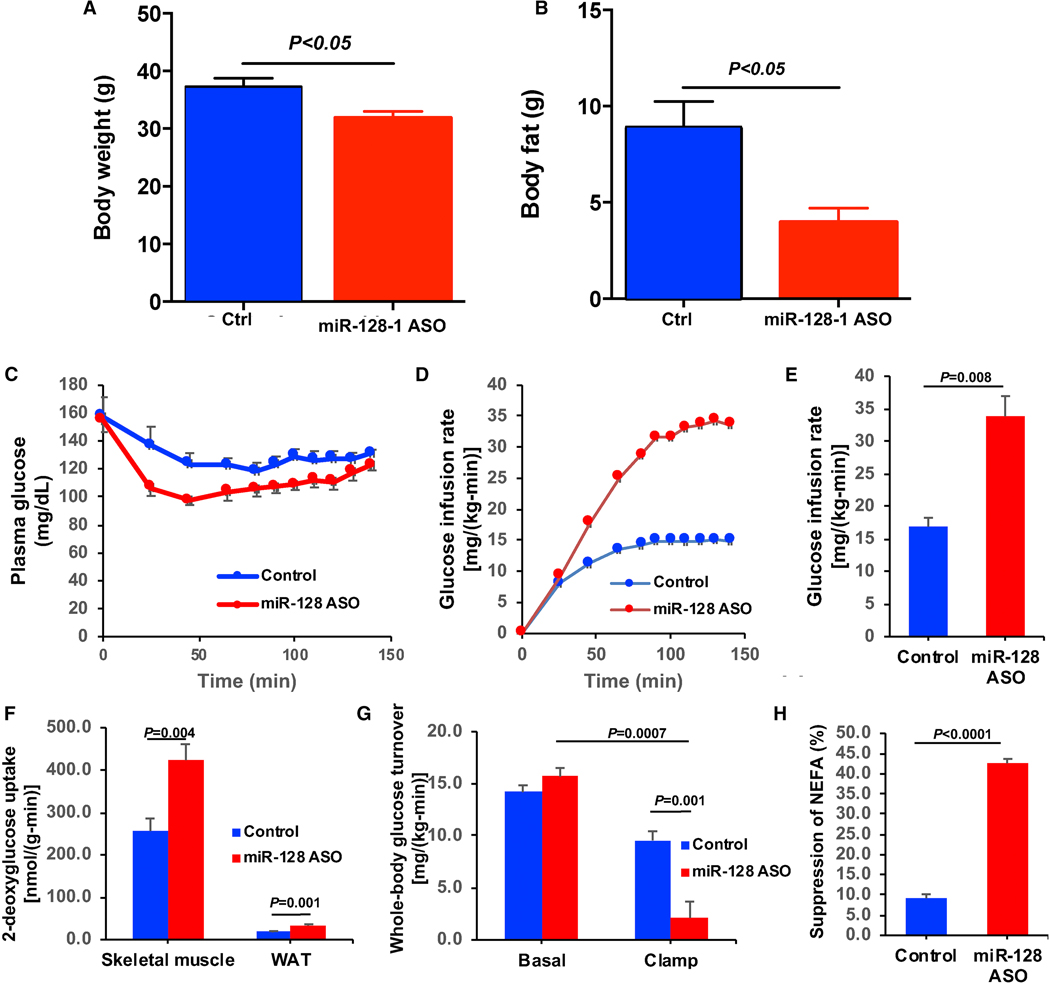
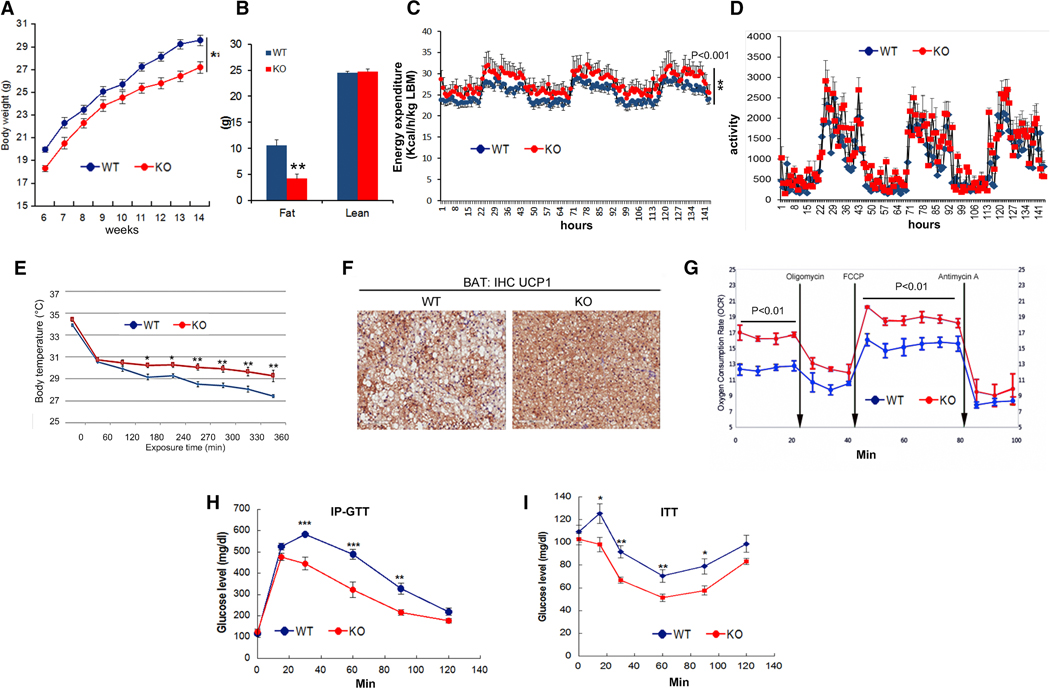
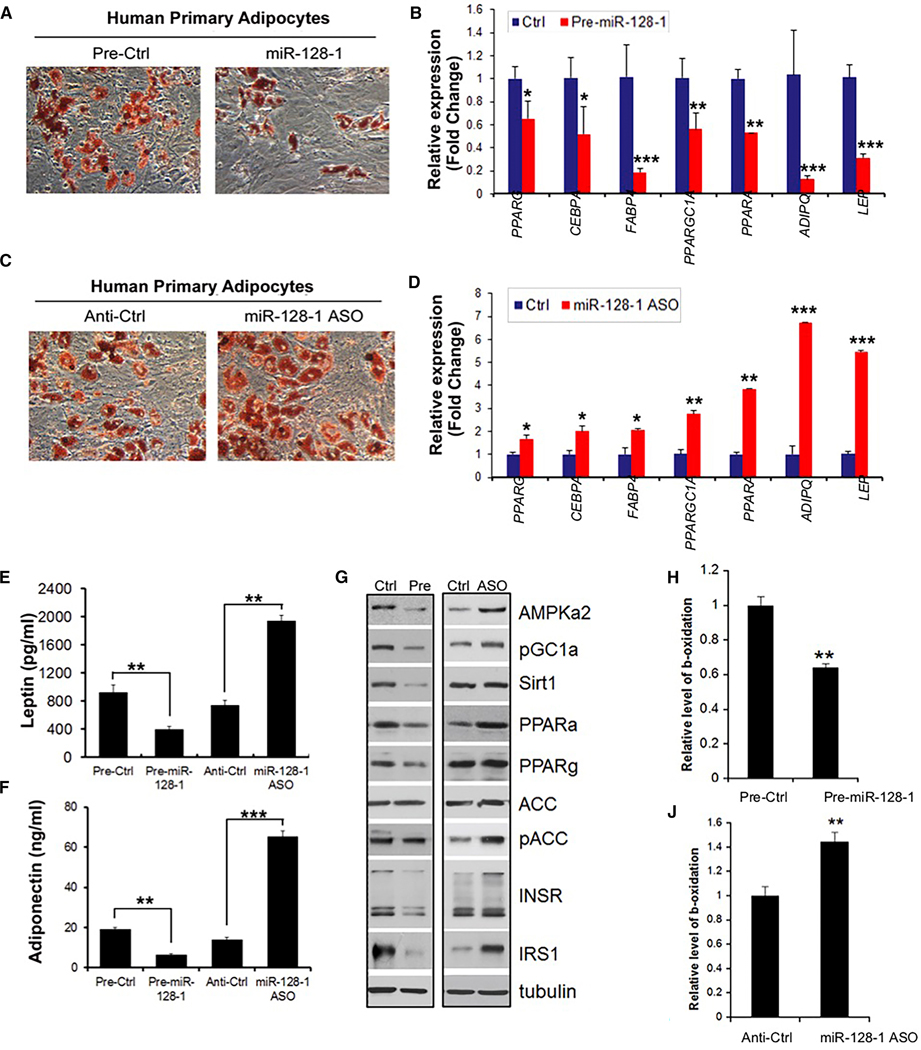
Positive selection in Europeans at the 2q21.3 locus harboring the lactase gene has been attributed to selection for the ability of adults to digest milk to survive famine in ancient times. However, the 2q21.3 locus is also associated with obesity and type 2 diabetes in humans, raising the possibility that additional genetic elements in the locus may have contributed to evolutionary adaptation to famine by promoting energy storage, but which now confer susceptibility to metabolic diseases. We show here that the miR-128-1 microRNA, located at the center of the positively selected locus, represents a crucial metabolic regulator in mammals. Antisense targeting and genetic ablation of miR-128-1 in mouse metabolic disease models result in increased energy expenditure and amelioration of high-fat-diet-induced obesity and markedly improved glucose tolerance. A thrifty phenotype connected to miR-128-1-dependent energy storage may link ancient adaptation to famine and modern metabolic maladaptation associated with nutritional overabundance.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests A.M.N. has issued patents on miR-128-1 (U.S. Pat. Nos. 9,045,749; 9,476,046; 9,789,132).
Figures



Comment in
-
A Selective Sweep Conceals a MicroRNA with Broad Metabolic Effects.Cell Metab. 2020 Nov 3;32(5):697-698. doi: 10.1016/j.cmet.2020.10.014. Cell Metab. 2020. PMID: 33147481
References
-
- Albuquerque D, Nobrega C, and Manco L. (2013). The lactase persistence −13910C>T polymorphism shows indication of association with abdominal obesity among Portuguese children. Acta paediatrica 102, e153–157. - PubMed
-
- Allison DB, Kaprio J, Korkeila M, Koskenvuo M, Neale MC, and Hayakawa K. (1996). The heritability of body mass index among an international sample of monozygotic twins reared apart. Int J Obes Relat Metab Disord 20, 501–6. - PubMed
-
- Ambros V. (2004). The functions of animal microRNAs. Nature 431, 350–355. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P30 DK040561/DK/NIDDK NIH HHS/United States
- P01 DA047233/DA/NIDA NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- R01 DK114793/DK/NIDDK NIH HHS/United States
- R01 DK116774/DK/NIDDK NIH HHS/United States
- R01 DK094184/DK/NIDDK NIH HHS/United States
- K99 HG008179/HG/NHGRI NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- R01 NS091574/NS/NINDS NIH HHS/United States
- R01 DK119968/DK/NIDDK NIH HHS/United States
- P30 DK046200/DK/NIDDK NIH HHS/United States
- R01 HD032443/HD/NICHD NIH HHS/United States
- R01 DK114277/DK/NIDDK NIH HHS/United States
- P30 DK057521/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
