

Integrated Neurophotonics: Toward Dense Volumetric Interrogation of Brain Circuit Activity-at Depth and in Real Time
- PMID: 33058767
- PMCID: PMC8061790
- DOI: 10.1016/j.neuron.2020.09.043
Integrated Neurophotonics: Toward Dense Volumetric Interrogation of Brain Circuit Activity-at Depth and in Real Time
Abstract
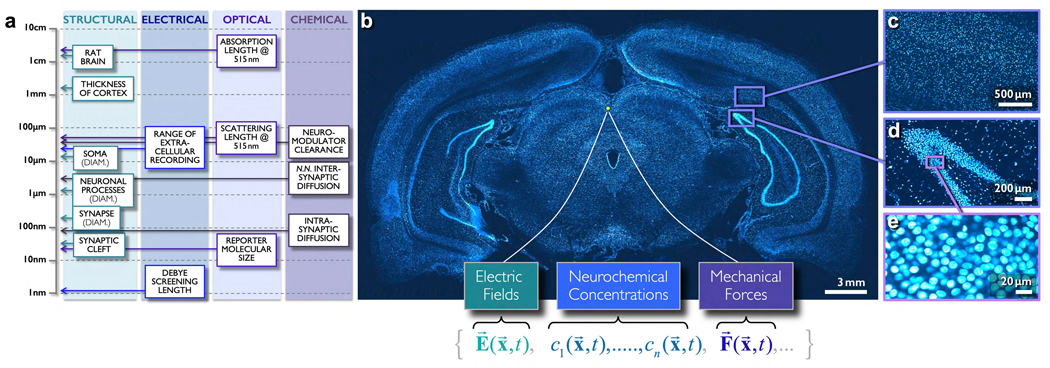
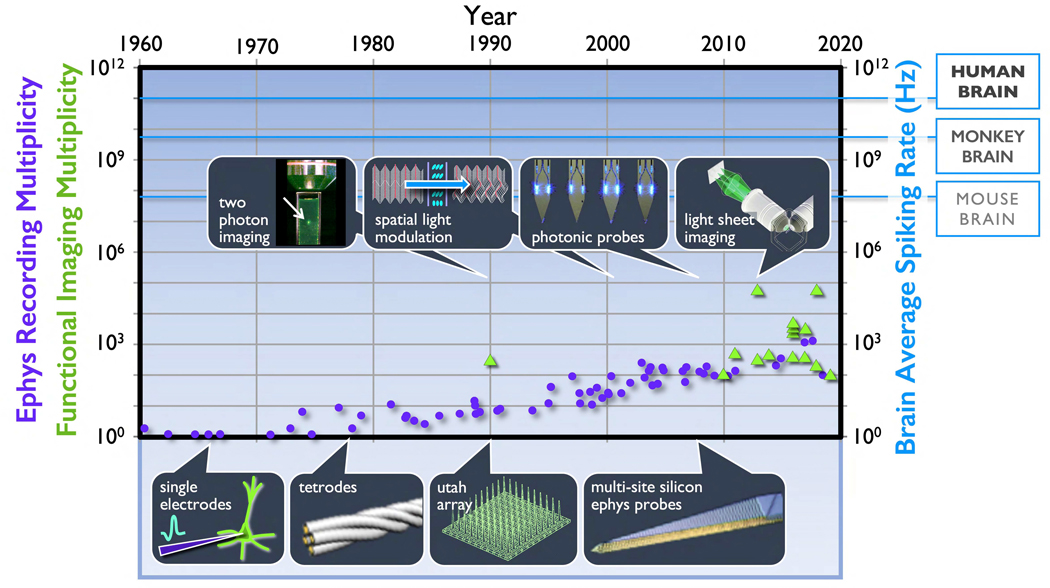
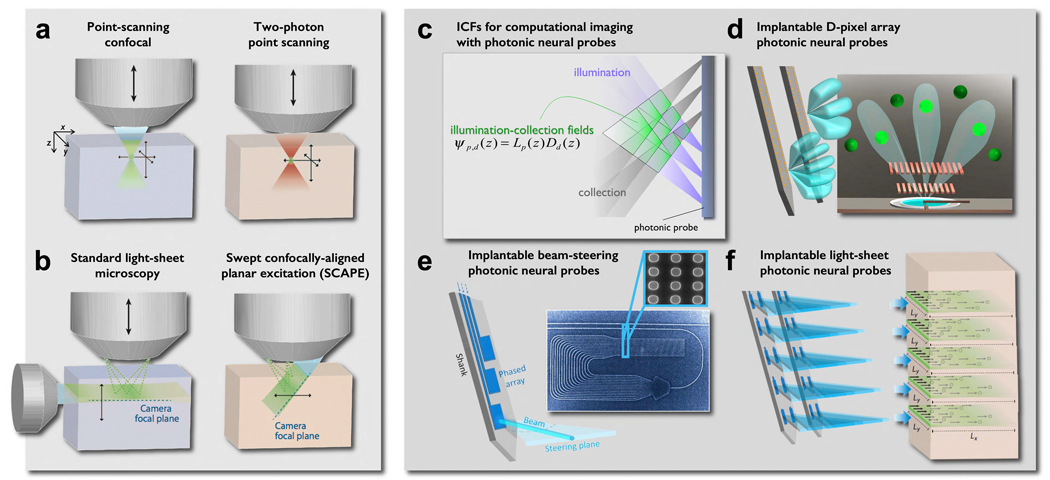
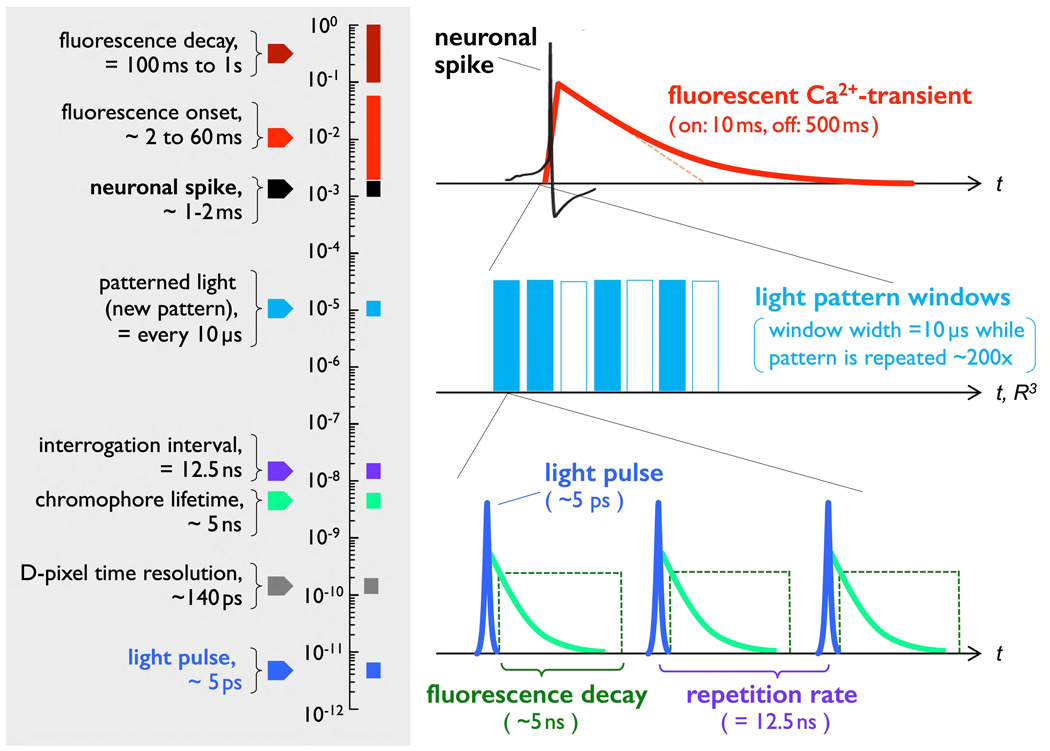
We propose a new paradigm for dense functional imaging of brain activity to surmount the limitations of present methodologies. We term this approach "integrated neurophotonics"; it combines recent advances in microchip-based integrated photonic and electronic circuitry with those from optogenetics. This approach has the potential to enable lens-less functional imaging from within the brain itself to achieve dense, large-scale stimulation and recording of brain activity with cellular resolution at arbitrary depths. We perform a computational study of several prototype 3D architectures for implantable probe-array modules that are designed to provide fast and dense single-cell resolution (e.g., within a 1-mm3 volume of mouse cortex comprising ∼100,000 neurons). We describe progress toward realizing integrated neurophotonic imaging modules, which can be produced en masse with current semiconductor foundry protocols for chip manufacturing. Implantation of multiple modules can cover extended brain regions.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests Three patents owned by the California Institute of Technology have been pursued in connection with this work: (1) Roukes (2011). Brain-machine interface based on photonic neural probe arrays. U.S.P.T.O. Patent No. 10,638,933, Issued May 5, 2020, Priority Date: December 8, 2011. (2) Segev, E., Moreaux, L.C., Fowler, T.M. Faraon, A., Roukes, M.L. Implantable, highly collimated light-emitters for biological applications. U.S.P.T.O. Patent No. 10,471,273, Issue date: 12 November 2019; Priority date: 16 October 2015. (3) Roukes et al. (2016). One-photon integrated neurophotonic systems. U.S.P.T.O. Patent Application 20160150963, Filing date: November 5, 2014; Publication date: June 2, 2016.
Figures





References
-
- Acher FC, and Bertrand HO (2005). Amino acid recognition by Venus flytrap domains is encoded in an 8-residue motif. Biopolymers 80, 357–366. - PubMed
-
- Ahrens MB, Orger MB, Robson DN, Li JM, and Keller PJ (2013). Whole-brain functional imaging at cellular resolution using light-sheet microscopy. Nature Methods 10, 413–420. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
