

Secondary ossification center induces and protects growth plate structure
- PMID: 33063669
- PMCID: PMC7581430
- DOI: 10.7554/eLife.55212
Secondary ossification center induces and protects growth plate structure
Abstract
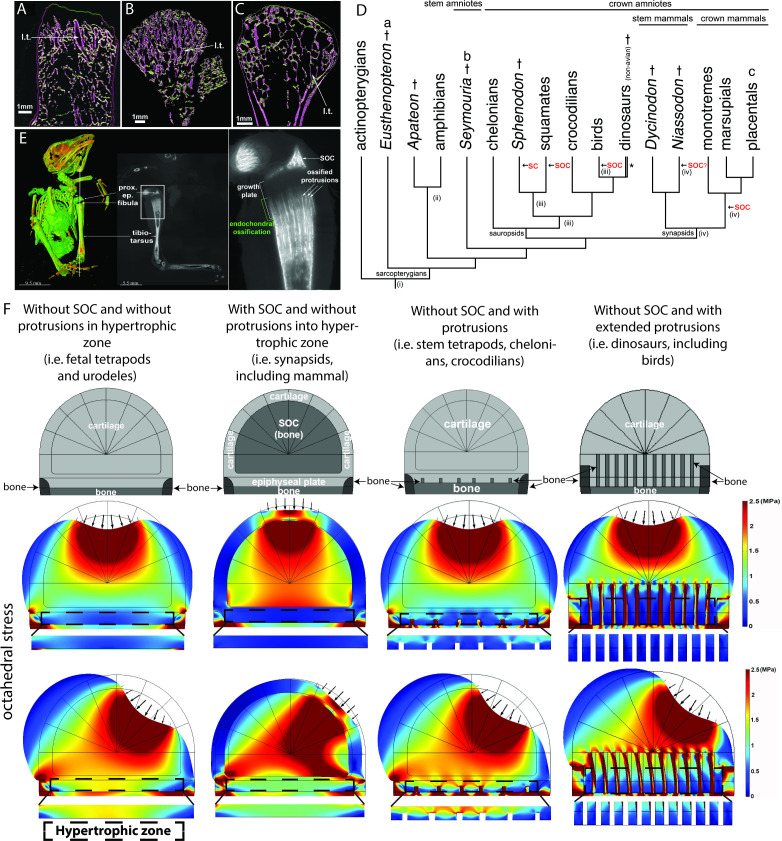
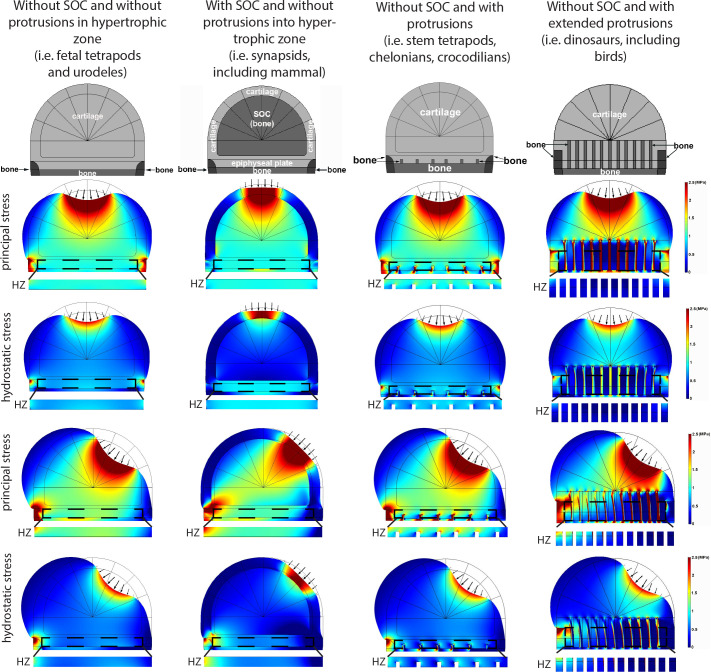
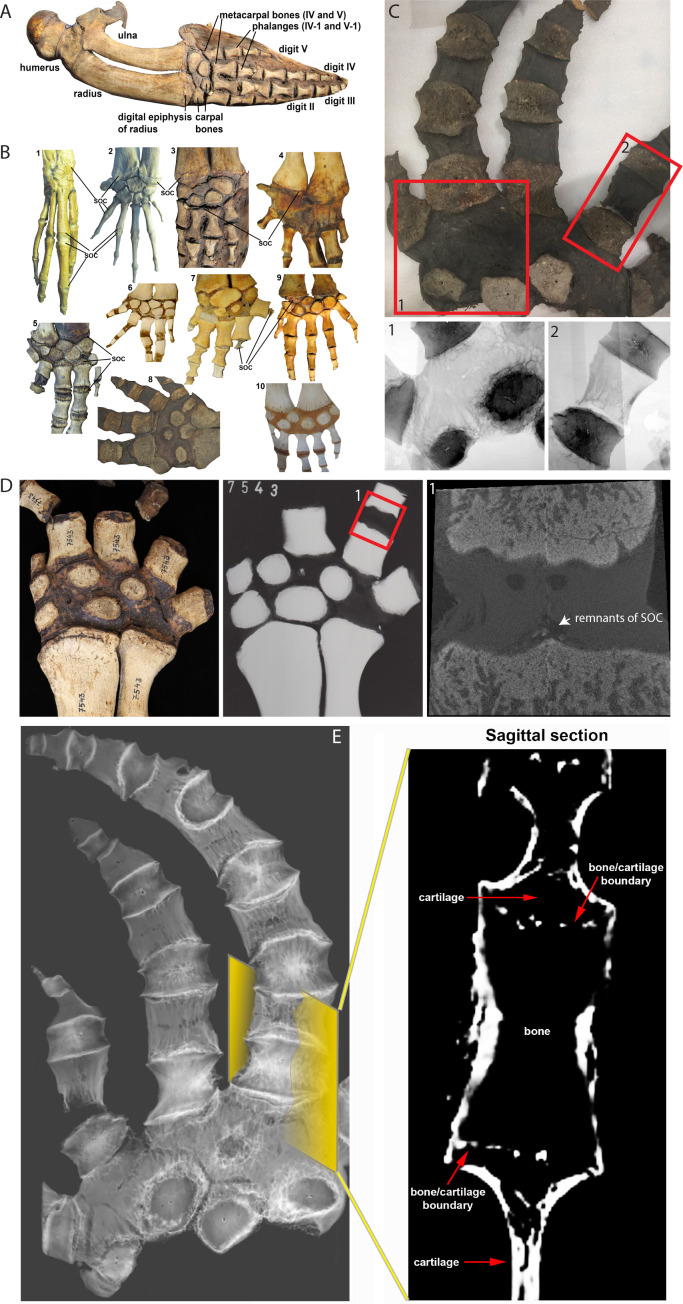
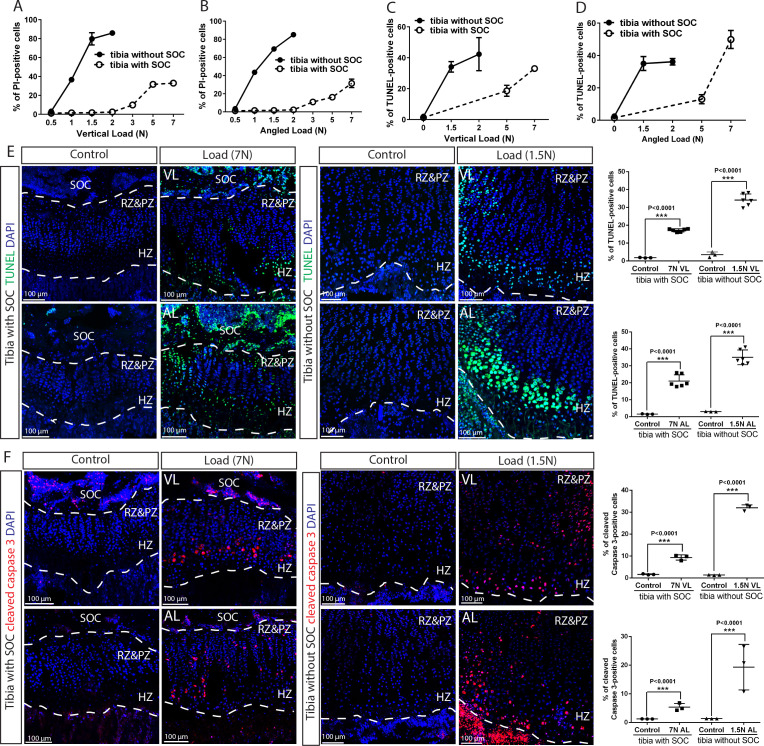
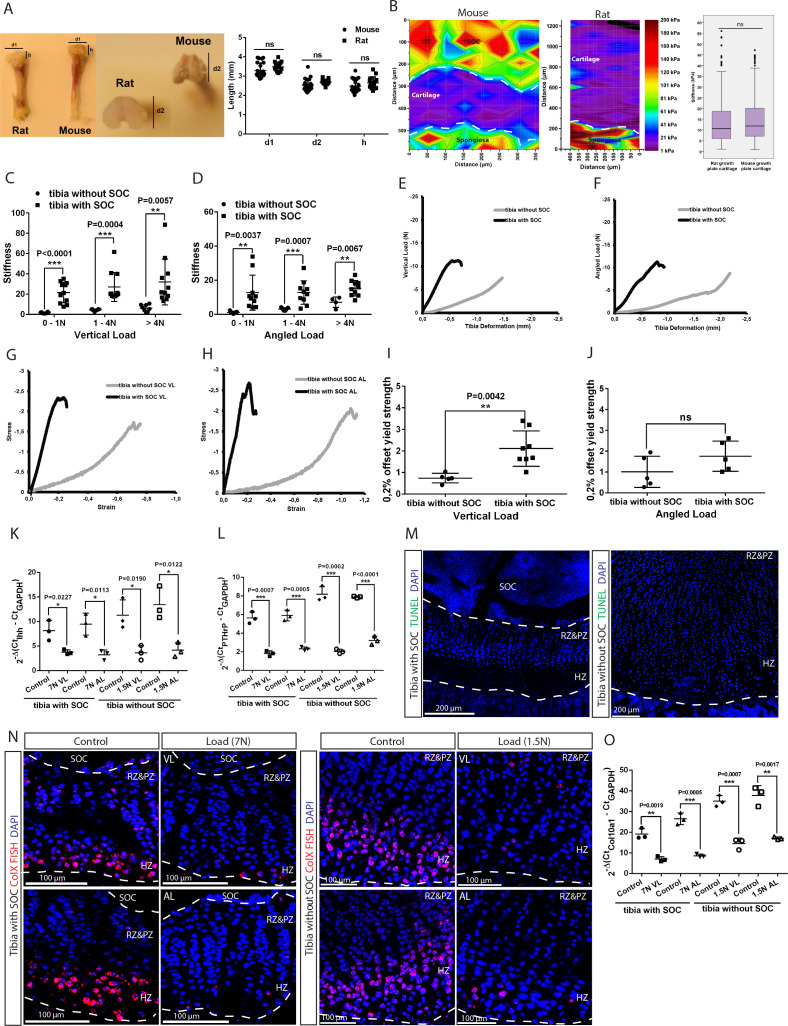
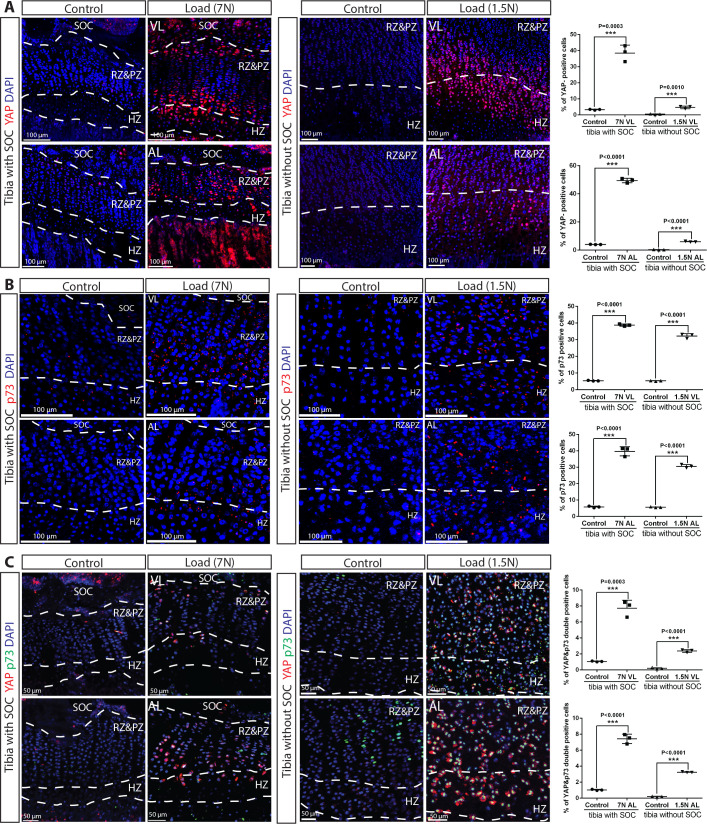
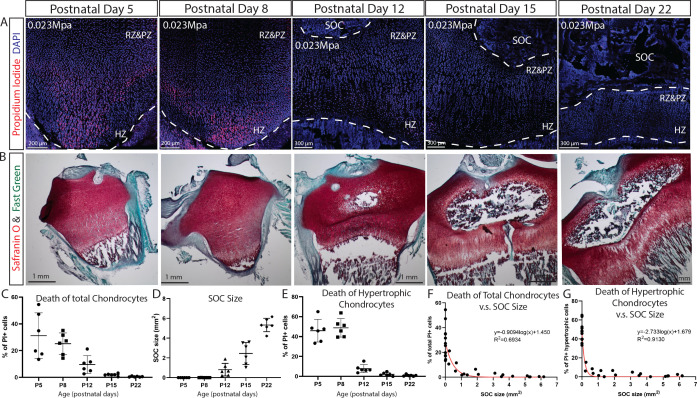
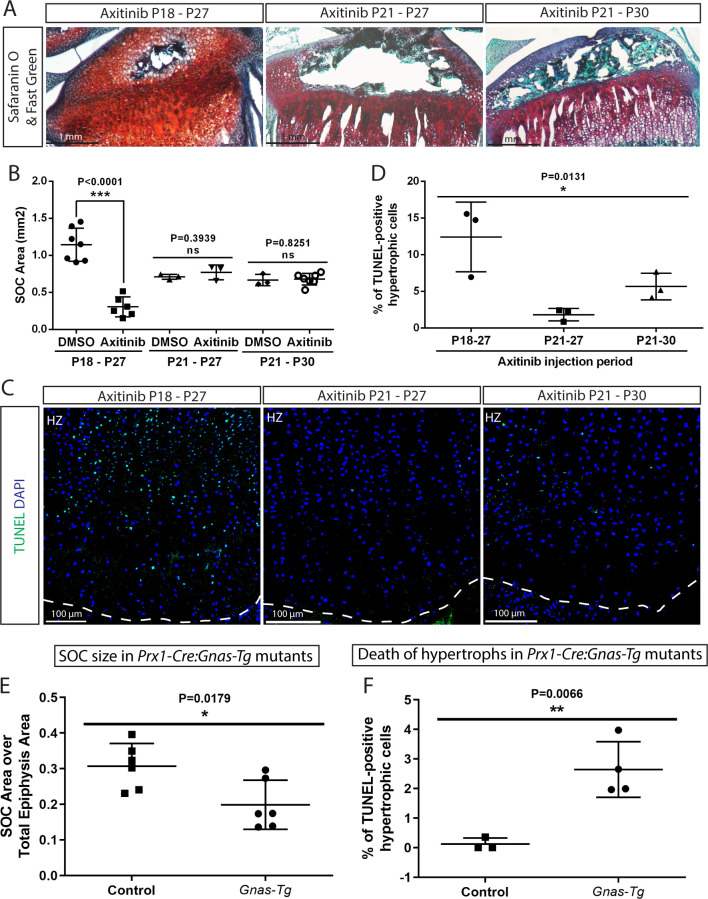
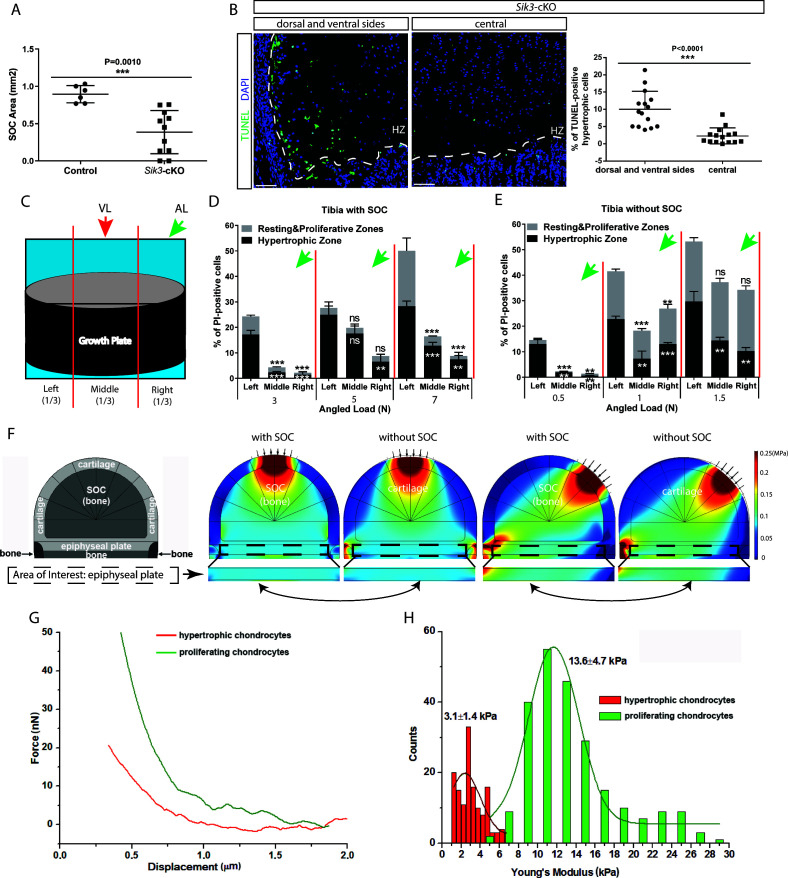
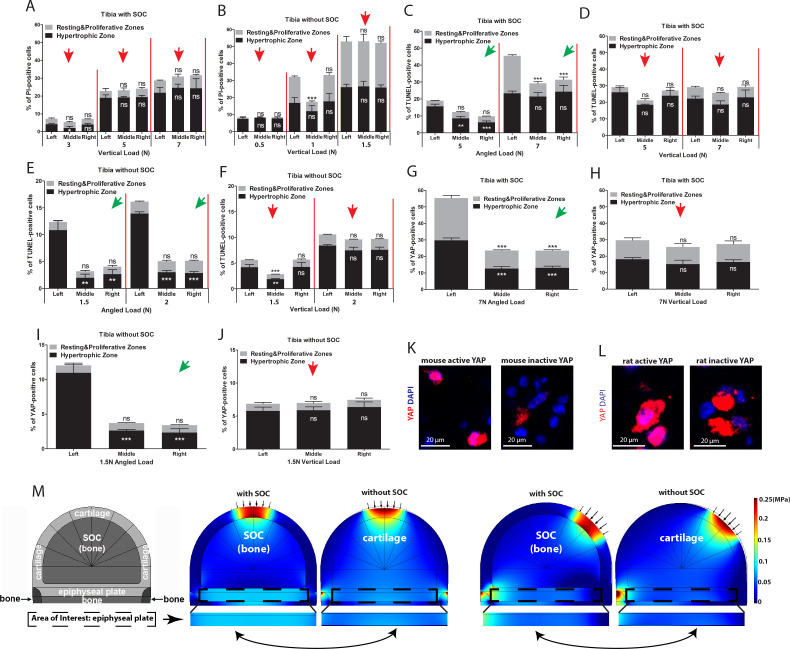
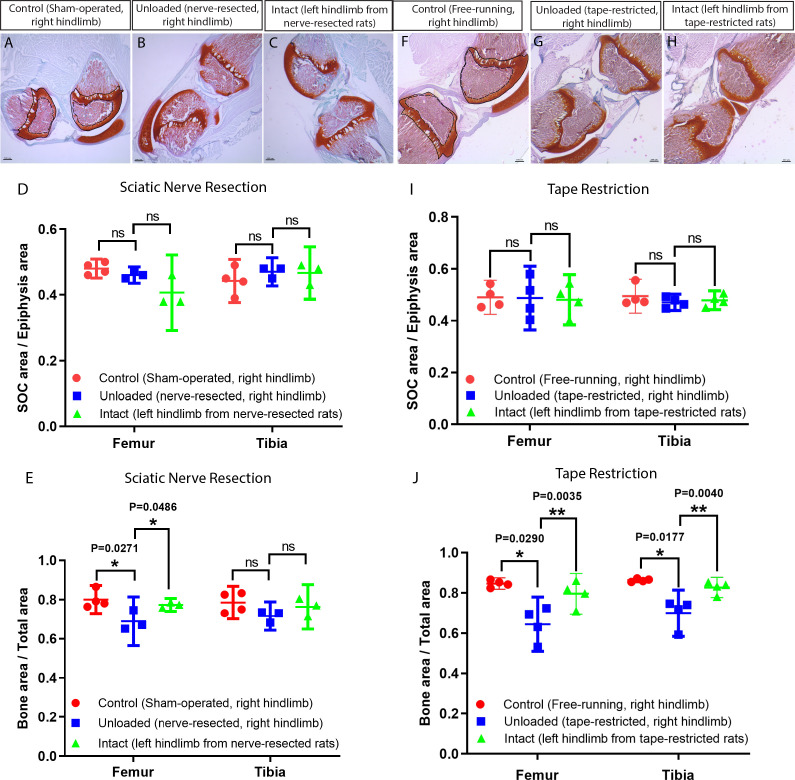
Growth plate and articular cartilage constitute a single anatomical entity early in development but later separate into two distinct structures by the secondary ossification center (SOC). The reason for such separation remains unknown. We found that evolutionarily SOC appears in animals conquering the land - amniotes. Analysis of the ossification pattern in mammals with specialized extremities (whales, bats, jerboa) revealed that SOC development correlates with the extent of mechanical loads. Mathematical modeling revealed that SOC reduces mechanical stress within the growth plate. Functional experiments revealed the high vulnerability of hypertrophic chondrocytes to mechanical stress and showed that SOC protects these cells from apoptosis caused by extensive loading. Atomic force microscopy showed that hypertrophic chondrocytes are the least mechanically stiff cells within the growth plate. Altogether, these findings suggest that SOC has evolved to protect the hypertrophic chondrocytes from the high mechanical stress encountered in the terrestrial environment.
Keywords: bat; chondrocytes; developmental biology; evolutionary biology; growth plate; jerboa; mouse; rat; secondary ossification; whale.
© 2020, Xie et al.
Conflict of interest statement
MX, PG, AH, JE, EM, LL, PN, SK, BS, AS, LS, BM, KG, SN, AA, AU, AK, AK, IA, PT, KF, MC, AS, CG, HK, MB, KC, PT, SS, IA, AE, AC No competing interests declared
Figures








References
-
- Adams RA, Thibault KM. Ontogeny and evolution of the hindlimb and calcar: assessing phylogenetic trends. Ontogeny, Functional Ecology, and Evolution of Bats. 2000;10:316–332. doi: 10.1017/CBO9780511541872.010. - DOI
-
- Alexander RM. Mechanics of posture and gait of some large dinosaurs. Zoological Journal of the Linnean Society. 1985;83:1–25. doi: 10.1111/j.1096-3642.1985.tb00871.x. - DOI
-
- Anderson JF, Hall-Martin A, Russell DA. Long-bone circumference and weight in mammals, birds and dinosaurs. Journal of Zoology. 1985;207:53–61. doi: 10.1111/j.1469-7998.1985.tb04915.x. - DOI
-
- Ashley-Ross M. Hindlimb kinematics during terrestrial locomotion in a salamander (Dicamptodon tenebrosus) The Journal of Experimental Biology. 1994;193:255–283. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
