

Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit During Gestation
- PMID: 33066440
- PMCID: PMC7589826
- DOI: 10.3390/ijms21207578
Profiling of Tryptophan Metabolic Pathways in the Rat Fetoplacental Unit During Gestation
Abstract
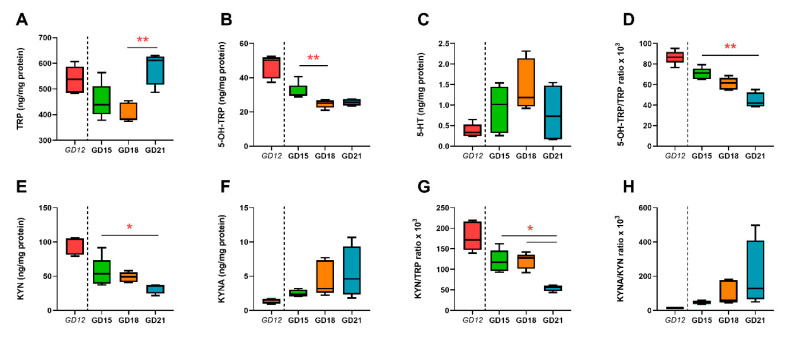
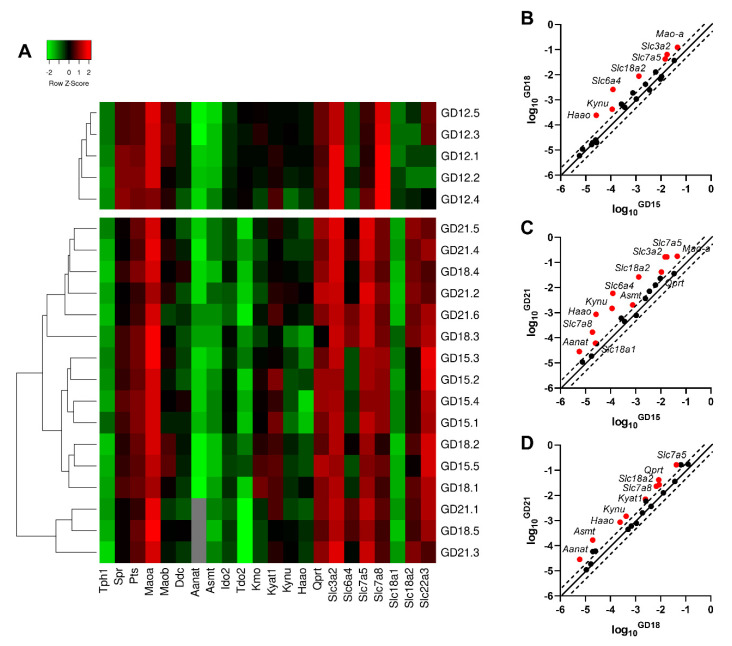
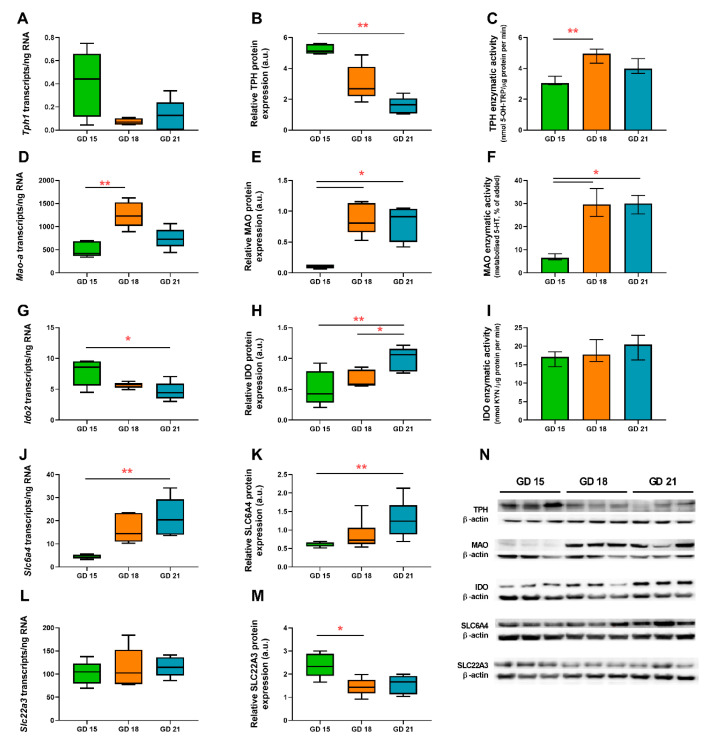
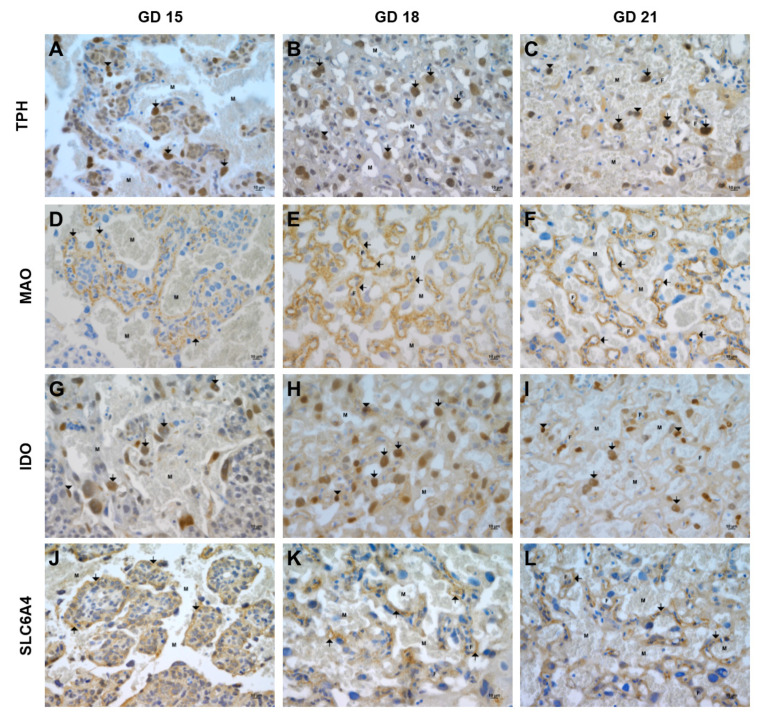
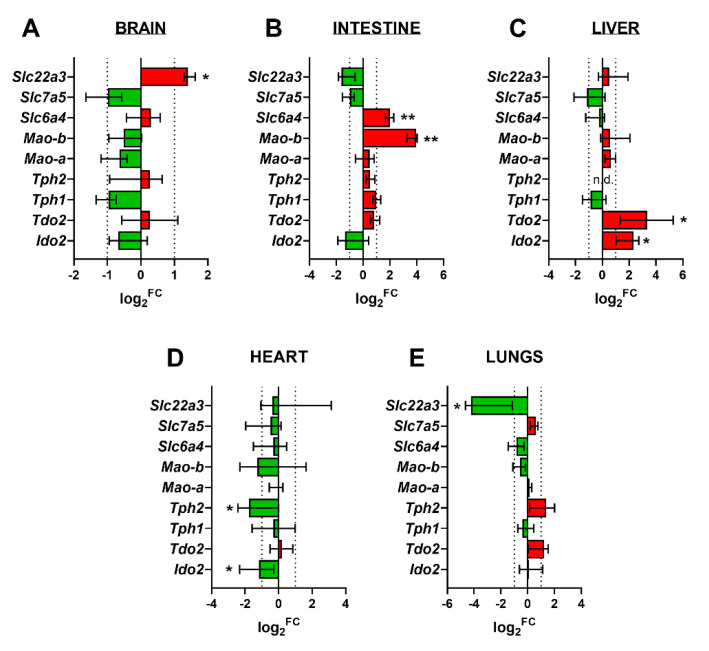
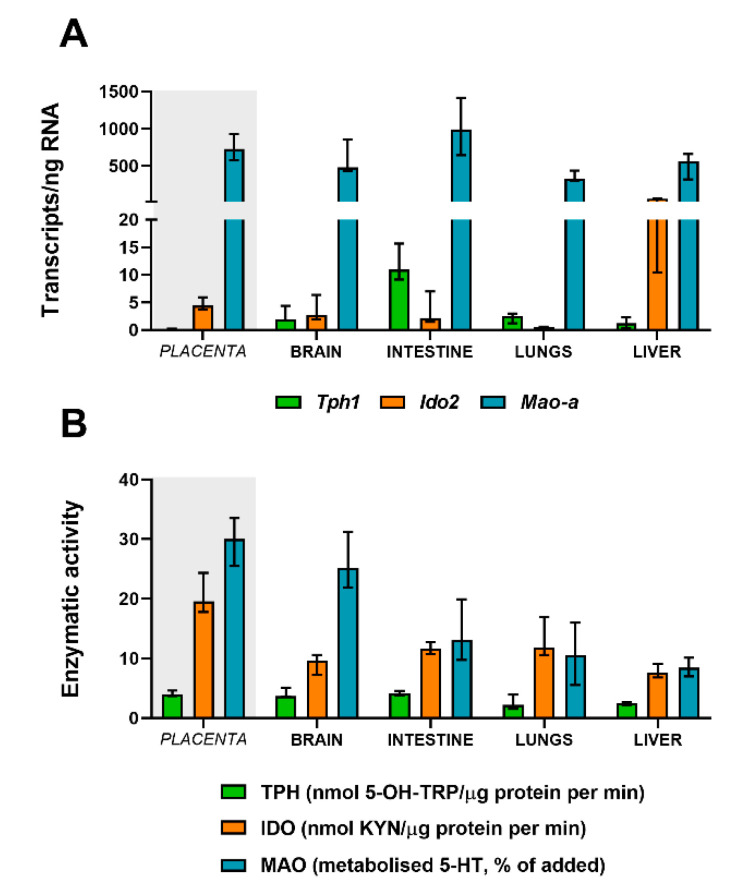
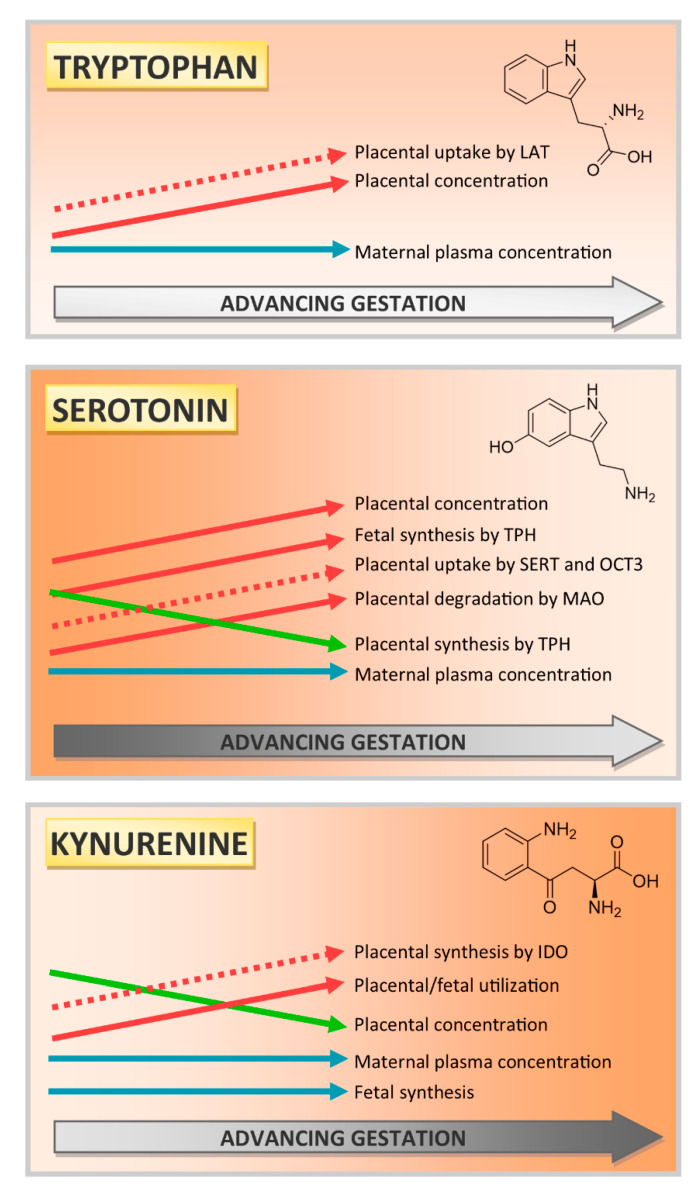
Placental homeostasis of tryptophan is essential for fetal development and programming. The two main metabolic pathways (serotonin and kynurenine) produce bioactive metabolites with immunosuppressive, neurotoxic, or neuroprotective properties and their concentrations in the fetoplacental unit must be tightly regulated throughout gestation. Here, we investigated the expression/function of key enzymes/transporters involved in tryptophan pathways during mid-to-late gestation in rat placenta and fetal organs. Quantitative PCR and heatmap analysis revealed the differential expression of several genes involved in serotonin and kynurenine pathways. To identify the flux of substrates through these pathways, Droplet Digital PCR, western blot, and functional analyses were carried out for the rate-limiting enzymes and transporters. Our findings show that placental tryptophan metabolism to serotonin is crucial in mid-gestation, with a subsequent switch to fetal serotonin synthesis. Concurrently, at term, the close interplay between transporters and metabolizing enzymes of both placenta and fetal organs orchestrates serotonin homeostasis and prevents hyper/hypo-serotonemia. On the other hand, the placental production of kynurenine increases during pregnancy, with a low contribution of fetal organs throughout gestation. Any external insult to this tightly regulated harmony of transporters and enzymes within the fetoplacental unit may affect optimal in utero conditions and have a negative impact on fetal programming.
Keywords: fetal organs; fetal programming; placenta–brain axis; pregnancy; rat model; tryptophan metabolism.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- Teshigawara T., Mouri A., Kubo H., Nakamura Y., Shiino T., Okada T., Morikawa M., Nabeshima T., Ozaki N., Yamamoto Y., et al. Changes in tryptophan metabolism during pregnancy and postpartum periods: Potential involvement in postpartum depressive symptoms. J. Affect. Disord. 2019;255:168–176. doi: 10.1016/j.jad.2019.05.028. - DOI - PubMed
-
- Zardoya-Laguardia P., Blaschitz A., Hirschmugl B., Lang I., A Herzog S., Nikitina L., Gauster M., Hausler M., Cervar-Zivkovic M., Karpf E., et al. Endothelial indoleamine 2,3-dioxygenase-1 regulates the placental vascular tone and is deficient in intrauterine growth restriction and pre-eclampsia. Sci. Rep. 2018;8:5488. doi: 10.1038/s41598-018-23896-0. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
