

The protein translation machinery is expressed for maximal efficiency in Escherichia coli
- PMID: 33067428
- PMCID: PMC7568582
- DOI: 10.1038/s41467-020-18948-x
The protein translation machinery is expressed for maximal efficiency in Escherichia coli
Abstract
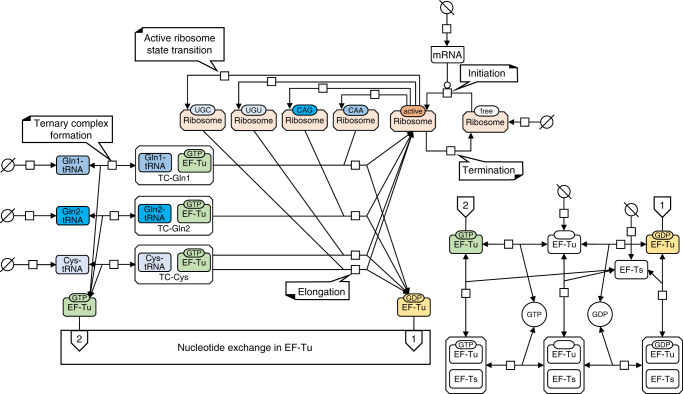
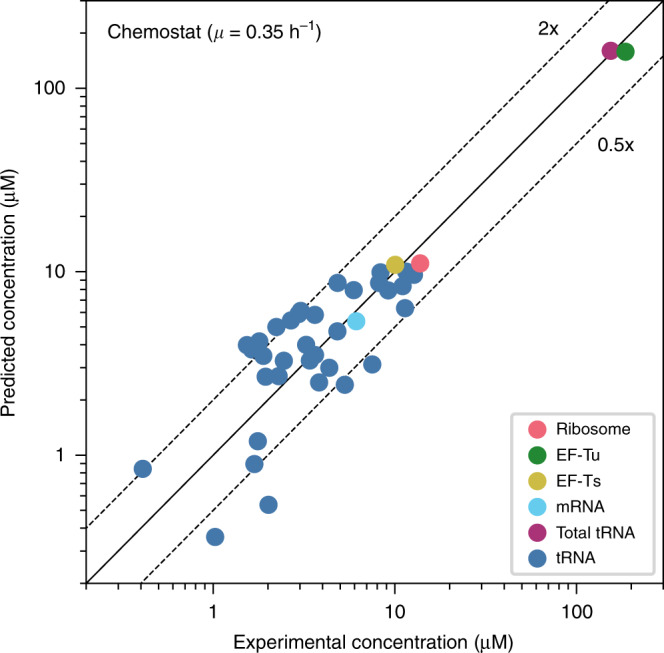
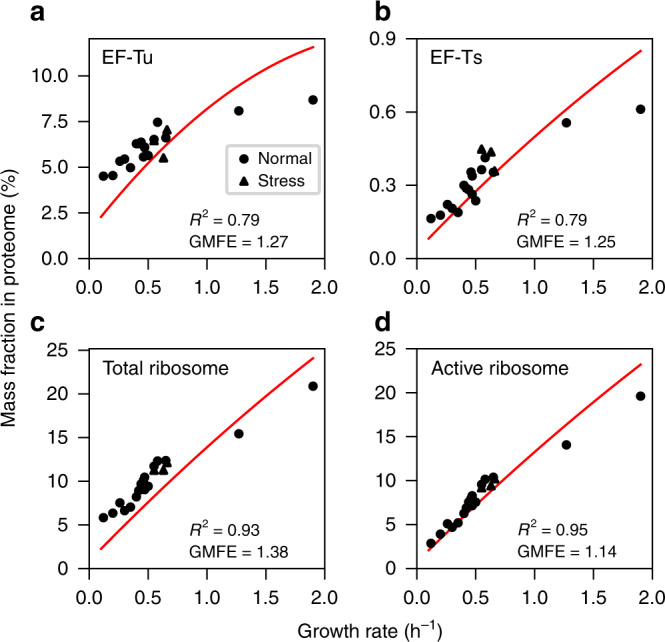
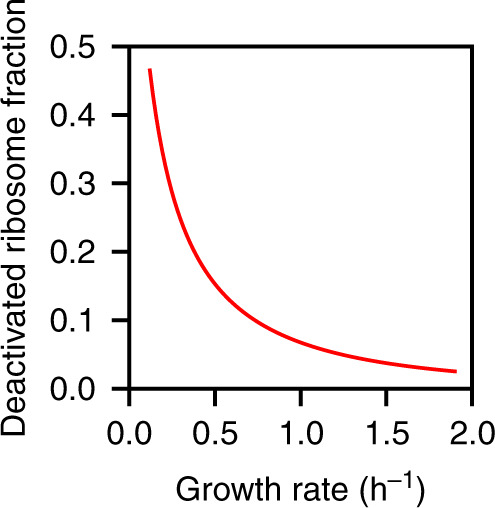
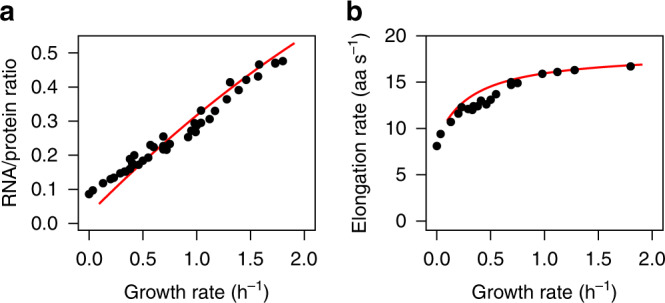
Protein synthesis is the most expensive process in fast-growing bacteria. Experimentally observed growth rate dependencies of the translation machinery form the basis of powerful phenomenological growth laws; however, a quantitative theory on the basis of biochemical and biophysical constraints is lacking. Here, we show that the growth rate-dependence of the concentrations of ribosomes, tRNAs, mRNA, and elongation factors observed in Escherichia coli can be predicted accurately from a minimization of cellular costs in a mechanistic model of protein translation. The model is constrained only by the physicochemical properties of the molecules and has no adjustable parameters. The costs of individual components (made of protein and RNA parts) can be approximated through molecular masses, which correlate strongly with alternative cost measures such as the molecules' carbon content or the requirement of energy or enzymes for their biosynthesis. Analogous cost minimization approaches may facilitate similar quantitative insights also for other cellular subsystems.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
[A model for trans-translation].Yi Chuan. 2006 Aug;28(8):1051-4. Yi Chuan. 2006. PMID: 16870596 Review. Chinese.
-
Mechanistic Modeling of In Vivo Translation in Escherichia coli Reliably Identifies Well-Adapted and Optimized RNA Sequences.ACS Synth Biol. 2025 Mar 21;14(3):699-710. doi: 10.1021/acssynbio.4c00578. Epub 2025 Feb 27. ACS Synth Biol. 2025. PMID: 40014843
-
Protein Synthesis in E. coli: Dependence of Codon-Specific Elongation on tRNA Concentration and Codon Usage.PLoS One. 2015 Aug 13;10(8):e0134994. doi: 10.1371/journal.pone.0134994. eCollection 2015. PLoS One. 2015. PMID: 26270805 Free PMC article.
-
Structural Basis for Polyproline-Mediated Ribosome Stalling and Rescue by the Translation Elongation Factor EF-P.Mol Cell. 2017 Nov 2;68(3):515-527.e6. doi: 10.1016/j.molcel.2017.10.014. Mol Cell. 2017. PMID: 29100052
-
tRNA engineering for manipulating genetic code.RNA Biol. 2018;15(4-5):453-460. doi: 10.1080/15476286.2017.1343227. Epub 2017 Sep 6. RNA Biol. 2018. PMID: 28722545 Free PMC article. Review.
Cited by
-
Structural Dynamics of RNA in the Presence of Choline Amino Acid Based Ionic Liquid: A Spectroscopic and Computational Outlook.ACS Cent Sci. 2021 Oct 27;7(10):1688-1697. doi: 10.1021/acscentsci.1c00768. Epub 2021 Sep 15. ACS Cent Sci. 2021. PMID: 34729412 Free PMC article.
-
How energy determines spatial localisation and copy number of molecules in neurons.Nat Commun. 2025 Feb 7;16(1):1424. doi: 10.1038/s41467-025-56640-0. Nat Commun. 2025. PMID: 39915472 Free PMC article.
-
Regulation of multiple paralogs of a small subunit ribosomal protein in Francisella tularensis.bioRxiv [Preprint]. 2025 Jun 29:2025.06.29.662229. doi: 10.1101/2025.06.29.662229. bioRxiv. 2025. PMID: 40667052 Free PMC article. Preprint.
-
Single-cell data reveal heterogeneity of investment in ribosomes across a bacterial population.Nat Commun. 2025 Jan 2;16(1):285. doi: 10.1038/s41467-024-55394-5. Nat Commun. 2025. PMID: 39746998 Free PMC article.
-
Current trends in biopharmaceuticals production in Escherichia coli.Biotechnol Lett. 2022 Aug;44(8):917-931. doi: 10.1007/s10529-022-03276-5. Epub 2022 Jul 7. Biotechnol Lett. 2022. PMID: 35796852 Review.
References
-
- Bremer, H. & Dennis, P. P. Modulation of chemical composition and other parameters of the cell by growth rate. In Escherichia coli and Salmonella: Cellular and Molecular Biology 1553–1569 (ASM Press, Washington, DC, 1996).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
