

Phenotypic flexibility in heat production and heat loss in response to thermal and hydric acclimation in the zebra finch, a small arid-zone passerine
- PMID: 33070274
- PMCID: PMC7819915
- DOI: 10.1007/s00360-020-01322-0
Phenotypic flexibility in heat production and heat loss in response to thermal and hydric acclimation in the zebra finch, a small arid-zone passerine
Erratum in
-
Correction to: Phenotypic flexibility in heat production and heat loss in response to thermal and hydric acclimation in the zebra finch, a small arid-zone passerine.J Comp Physiol B. 2021 Jan;191(1):241. doi: 10.1007/s00360-020-01327-9. J Comp Physiol B. 2021. PMID: 33247769 Free PMC article. No abstract available.
Abstract
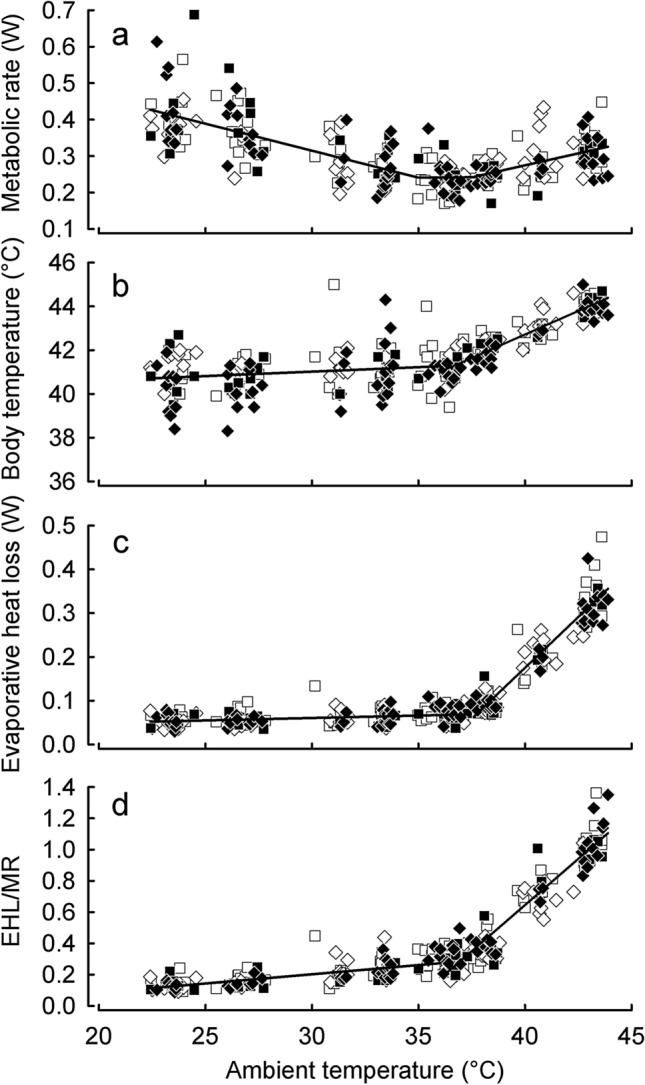
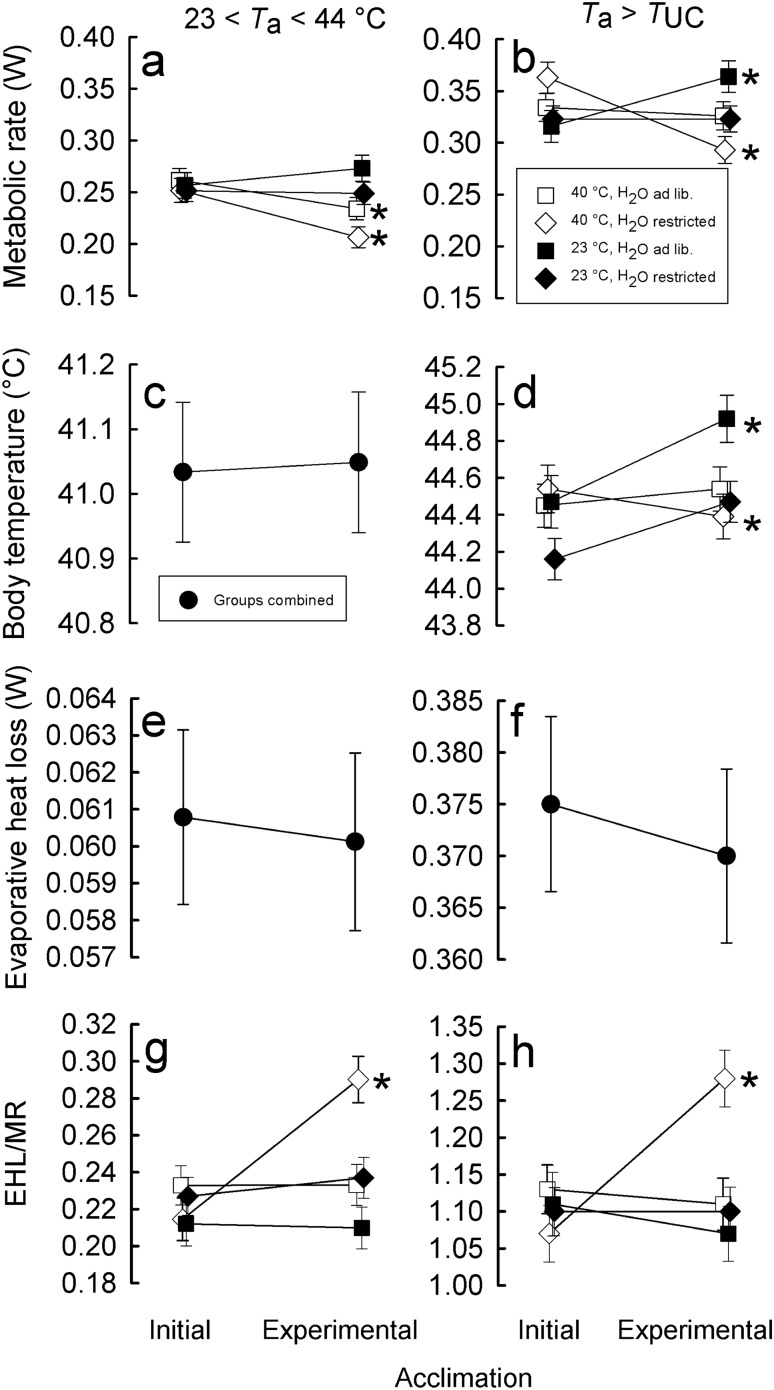
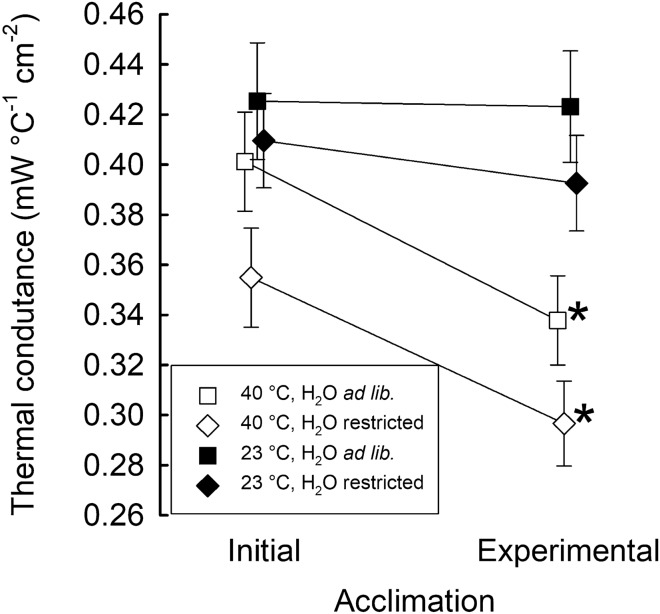
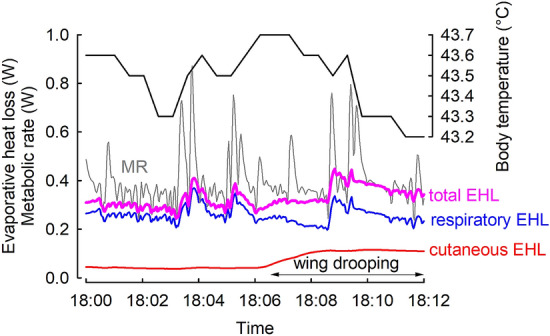
To maintain constant body temperature (Tb) over a wide range of ambient temperatures (Ta) endothermic animals require large amounts of energy and water. In hot environments, the main threat to endothermic homeotherms is insufficient water to supply that necessary for thermoregulation. We investigated flexible adjustment of traits related to thermoregulation and water conservation during acclimation to hot conditions or restricted water availability, or both, in the zebra finch, Taeniopygia guttata a small arid-zone passerine. Using indirect calorimetry, we measured changes in whole animal metabolic rate (MR), evaporative heat loss (EHL) and Tb before and after acclimation to 23 or 40 °C, with different availability of water. Additionally, we quantified changes in partitioning of EHL into respiratory and cutaneous avenues in birds exposed to 25 and 40 °C. In response to heat and water restriction zebra finches decreased MR, which together with unchanged EHL resulted in increased efficiency of evaporative heat loss. This facilitated more precise Tb regulation in heat-acclimated birds. Acclimation temperature and water availability had no effect on the partitioning of EHL into cutaneous or respiratory avenues. At 25 °C, cutaneous EHL accounted for ~ 60% of total EHL, while at 40 °C, its contribution decreased to ~ 20%. Consistent among-individual differences in MR and EHL suggest that these traits, provided that they are heritable, may be a subject to natural selection. We conclude that phenotypic flexibility in metabolic heat production associated with acclimation to hot, water-scarce conditions is crucial in response to changing environmental conditions, especially in the face of current and predicted climate change.
Keywords: Energy metabolism; Evaporative heat loss; Passerine; Phenotypic flexibility; Thermoregulation.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures


References
-
- Arad Z, Gavrieli-Levin I, Eylath U, Marder J. Effect of dehydration on cutaneous water evaporation in heat-exposed pigeons (Columba livia) Physiol Zool. 1987;60(6):623–630. doi: 10.1086/physzool.60.6.30159978. - DOI
-
- Aschoff J. Thermal conductance in mammals and birds: Its dependence on body size and crcadian phase. Comp Biochem Physiol A Physiol. 1981;69(4):611–619. doi: 10.1016/0300-9629(81)90145-6. - DOI
-
- Bakken GS. Measurement and application of operative and standard operative temperatures in ecology. Am Zool. 1992;32(2):194–216. doi: 10.1093/icb/32.2.194. - DOI
-
- Barcelo G, Love OP, Vezina F. Uncoupling basal and summit metabolic rates in white-throated sparrows: digestive demand drives maintenance costs, but changes in muscle mass are not needed to improve thermogenic capacity. Physiol Biochem Zool PBZ. 2017;90(2):153–165. doi: 10.1086/689290. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
