

Divergent genes in gerbils: prevalence, relation to GC-biased substitution, and phenotypic relevance
- PMID: 33076817
- PMCID: PMC7574485
- DOI: 10.1186/s12862-020-01696-3
Divergent genes in gerbils: prevalence, relation to GC-biased substitution, and phenotypic relevance
Abstract
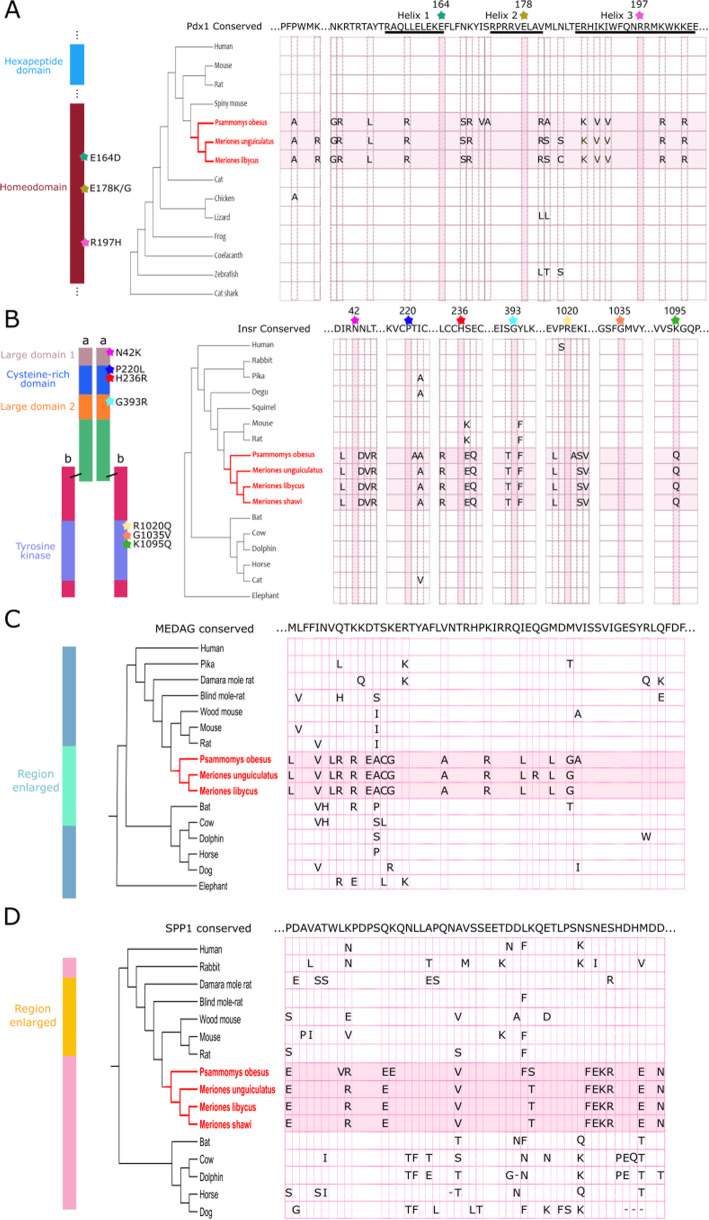
Background: Two gerbil species, sand rat (Psammomys obesus) and Mongolian jird (Meriones unguiculatus), can become obese and show signs of metabolic dysregulation when maintained on standard laboratory diets. The genetic basis of this phenotype is unknown. Recently, genome sequencing has uncovered very unusual regions of high guanine and cytosine (GC) content scattered across the sand rat genome, most likely generated by extreme and localized biased gene conversion. A key pancreatic transcription factor PDX1 is encoded by a gene in the most extreme GC-rich region, is remarkably divergent and exhibits altered biochemical properties. Here, we ask if gerbils have proteins in addition to PDX1 that are aberrantly divergent in amino acid sequence, whether they have also become divergent due to GC-biased nucleotide changes, and whether these proteins could plausibly be connected to metabolic dysfunction exhibited by gerbils.
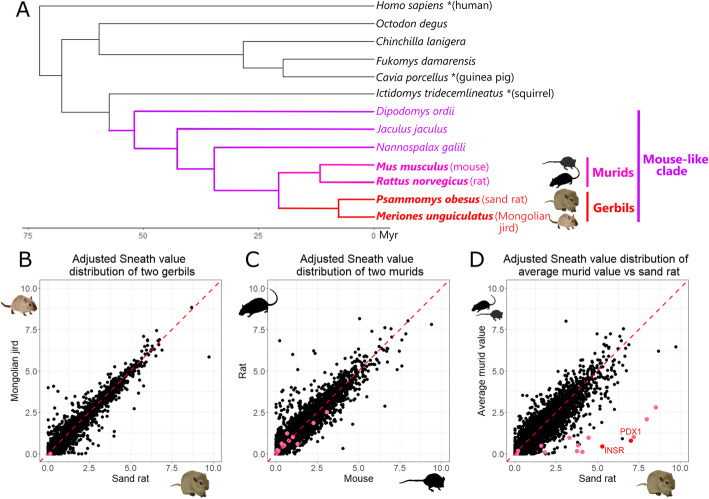
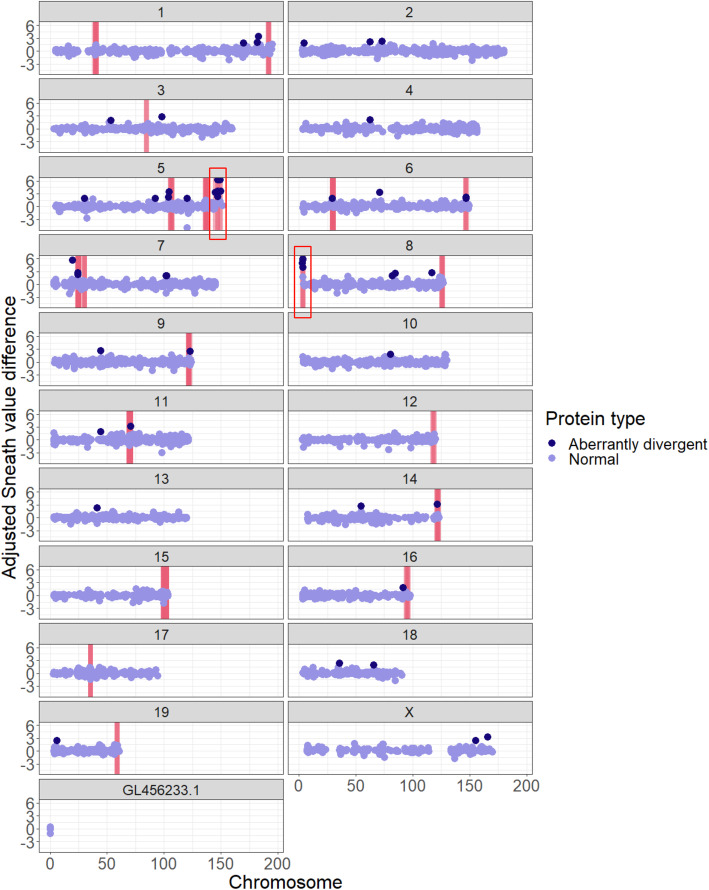
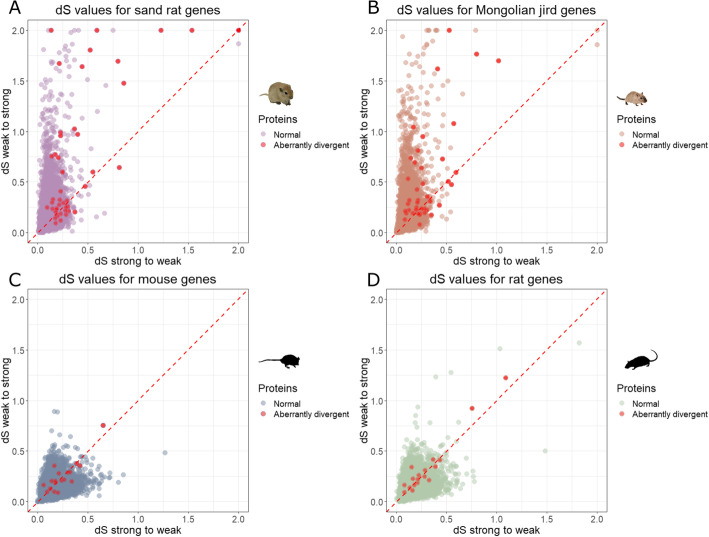
Results: We analyzed ~ 10,000 proteins with 1-to-1 orthologues in human and rodents and identified 50 proteins that accumulated unusually high levels of amino acid change in the sand rat and 41 in Mongolian jird. We show that more than half of the aberrantly divergent proteins are associated with GC biased nucleotide change and many are in previously defined high GC regions. We highlight four aberrantly divergent gerbil proteins, PDX1, INSR, MEDAG and SPP1, that may plausibly be associated with dietary metabolism.
Conclusions: We show that through the course of gerbil evolution, many aberrantly divergent proteins have accumulated in the gerbil lineage, and GC-biased nucleotide substitution rather than positive selection is the likely cause of extreme divergence in more than half of these. Some proteins carry putatively deleterious changes that could be associated with metabolic and physiological phenotypes observed in some gerbil species. We propose that these animals provide a useful model to study the 'tug-of-war' between natural selection and the excessive accumulation of deleterious substitutions mutations through biased gene conversion.
Keywords: GC bias; Genome evolution; Insulin receptor; Medag; Metabolism; Osteopontin; Pancreatic duodenal homeobox 1; Protein evolution; gBGC.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

References
-
- Leibowitz G, Ferber S, Apelqvist A, Edlund H, Gross DJ, Cerasi E, et al. IPF1/PDX1 deficiency and beta-cell dysfunction in Psammomys obesus, an animal with type 2 diabetes. Diabetes. 2001;50(8):1799–806. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
