

Sensory neurons from dorsal root ganglia regulate endothelial cell function in extracellular matrix remodelling
- PMID: 33076927
- PMCID: PMC7574530
- DOI: 10.1186/s12964-020-00656-0
Sensory neurons from dorsal root ganglia regulate endothelial cell function in extracellular matrix remodelling
Abstract
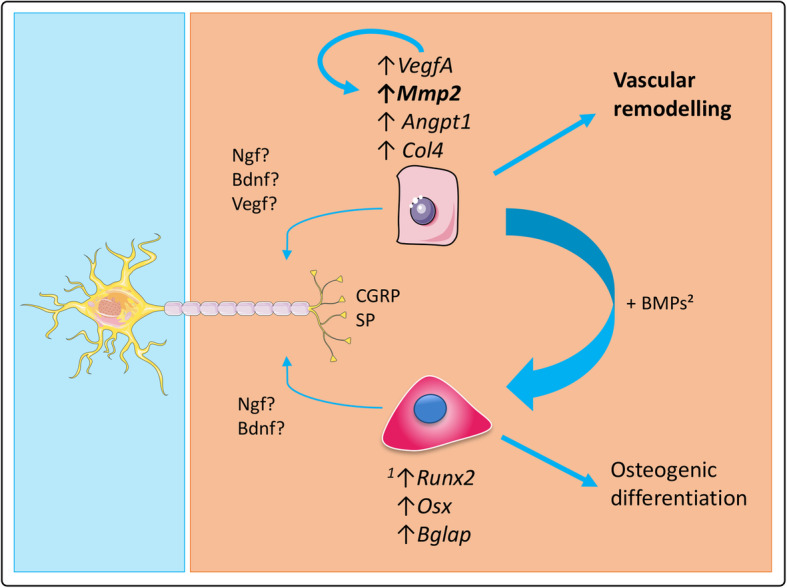
Background: Recent physiological and experimental data highlight the role of the sensory nervous system in bone repair, but its precise role on angiogenesis in a bone regeneration context is still unknown. Our previous work demonstrated that sensory neurons (SNs) induce the osteoblastic differentiation of mesenchymal stem cells, but the influence of SNs on endothelial cells (ECs) was not studied.
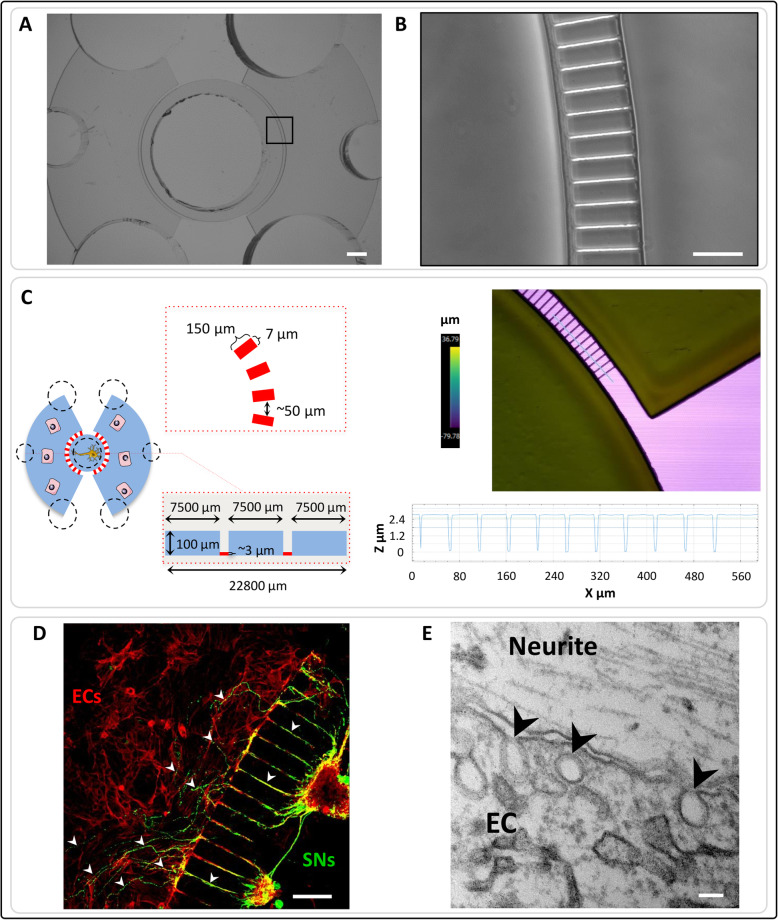
Methods: Here, in order to study in vitro the interplay between SNs and ECs, we used microfluidic devices as an indirect co-culture model. Gene expression analysis of angiogenic markers, as well as measurements of metalloproteinases protein levels and enzymatic activity, were performed.
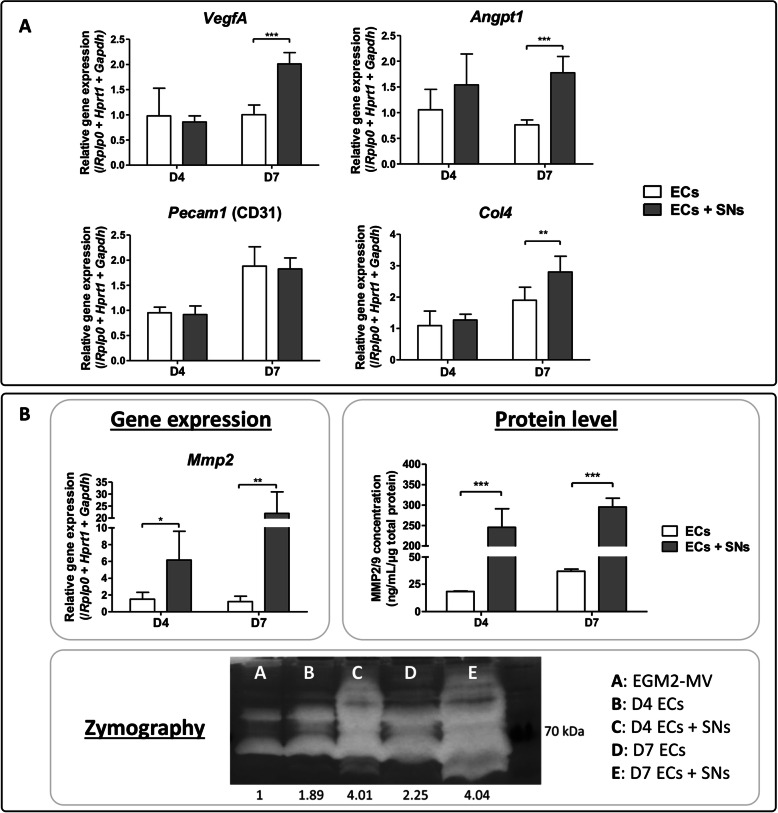
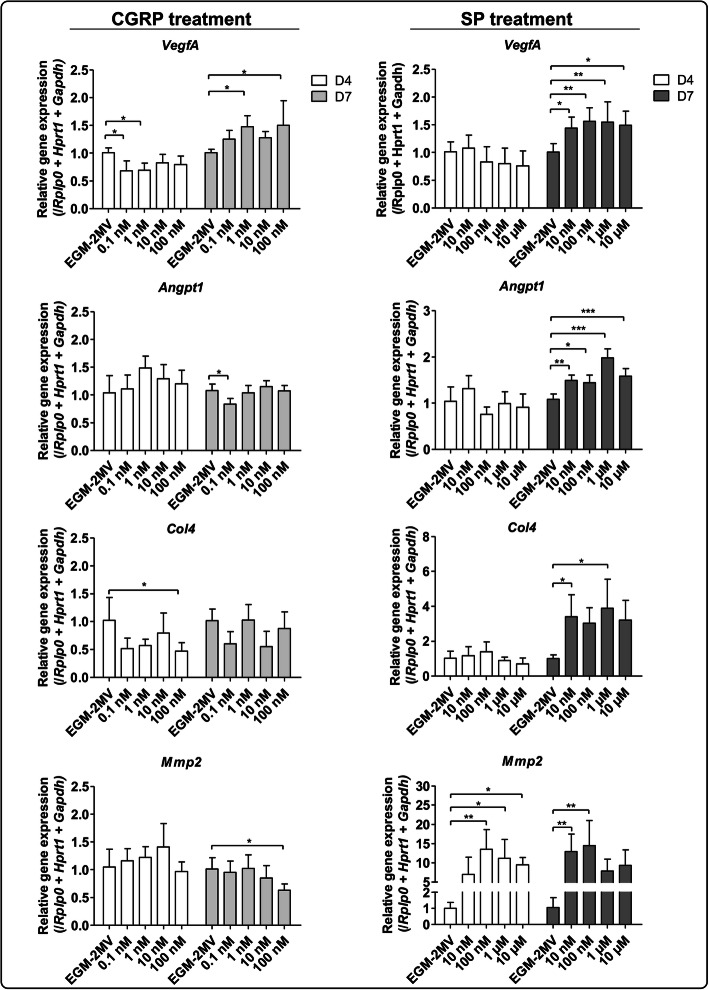
Results: We were able to demonstrate that two sensory neuropeptides, calcitonin gene-related peptide (CGRP) and substance P (SP), were involved in the transcriptional upregulation of angiogenic markers (vascular endothelial growth factor, angiopoietin 1, type 4 collagen, matrix metalloproteinase 2) in ECs. Co-cultures of ECs with SNs also increased the protein level and enzymatic activity of matrix metalloproteinases 2 and 9 (MMP2/MMP9) in ECs.
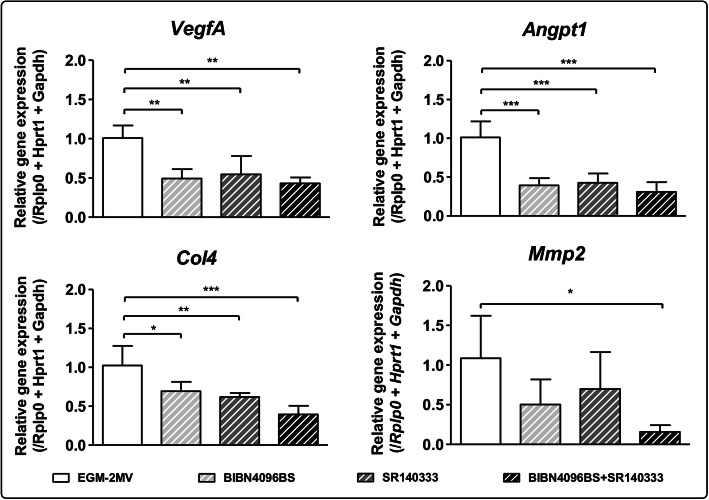
Conclusions: Our results suggest a role of sensory neurons, and more specifically of CGRP and SP, in the remodelling of endothelial cells extracellular matrix, thus supporting and enhancing the angiogenesis process. Video abstract.
Keywords: Angiogenesis; Cellular communication; Extracellular matrix Remodelling; Innervation; Matrix Metalloproteinases; Neurovascular interplay.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Miller MR, Kasahara M. Observations on the innervation of human long bones. Anat Rec. 1963;145(1):13–23. doi: 10.1002/ar.1091450104. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
