

Mosaic fungal individuals have the potential to evolve within a single generation
- PMID: 33077756
- PMCID: PMC7572425
- DOI: 10.1038/s41598-020-74679-5
Mosaic fungal individuals have the potential to evolve within a single generation
Abstract
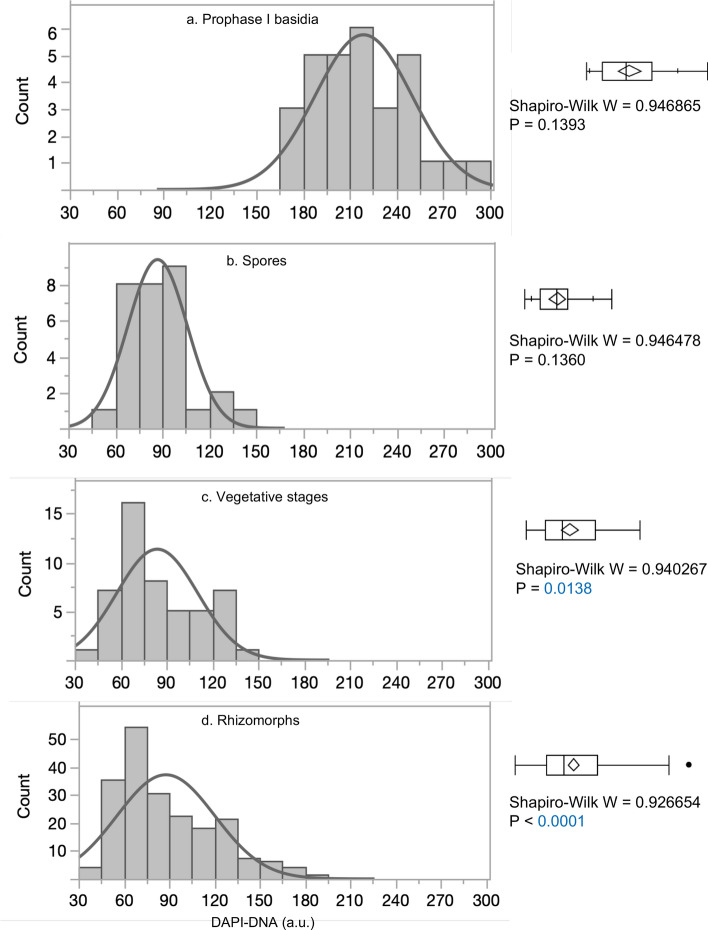
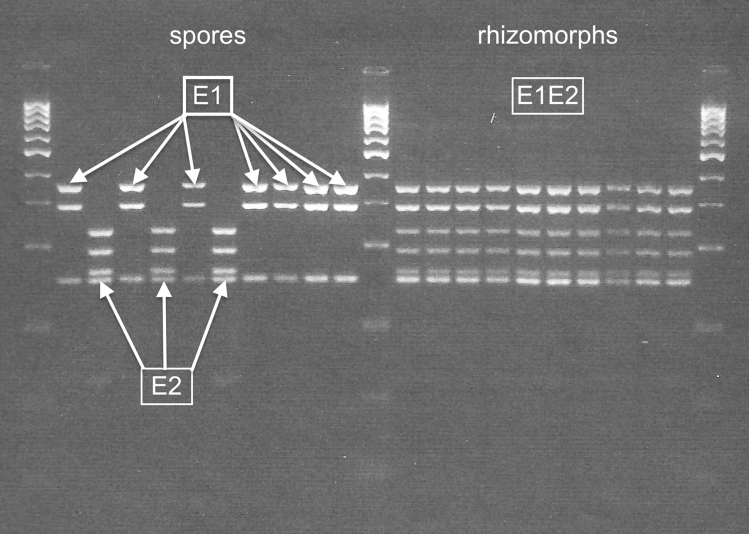
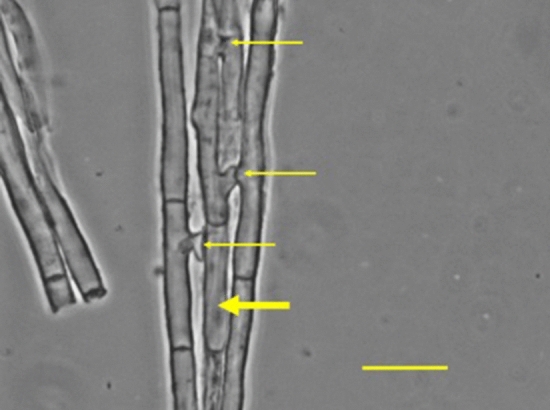
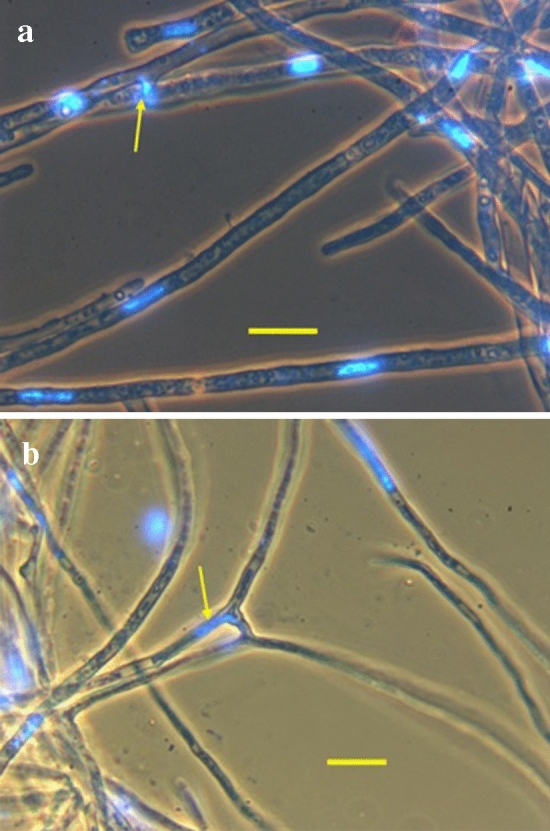
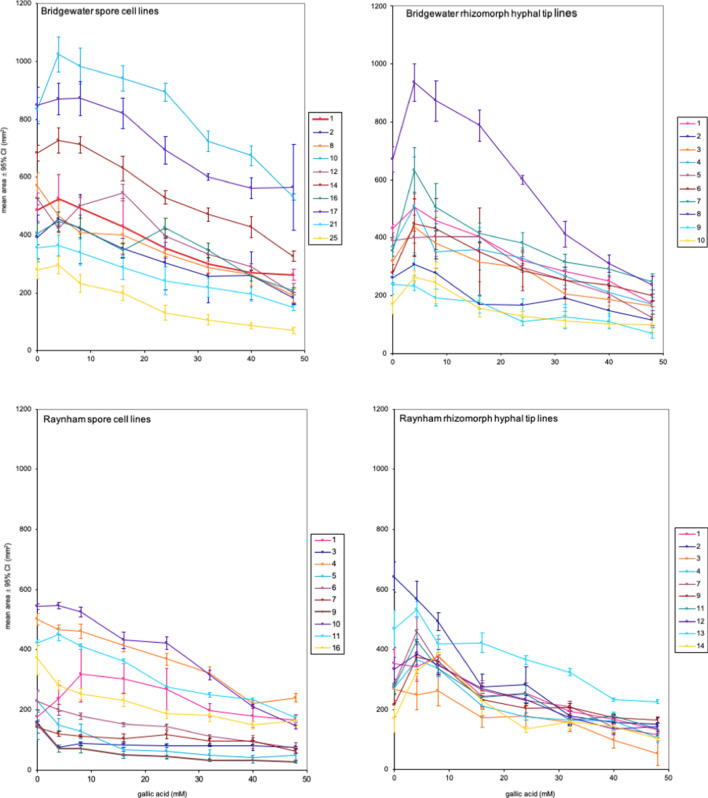
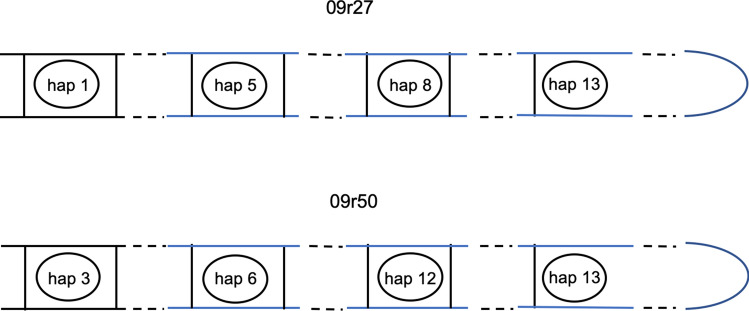
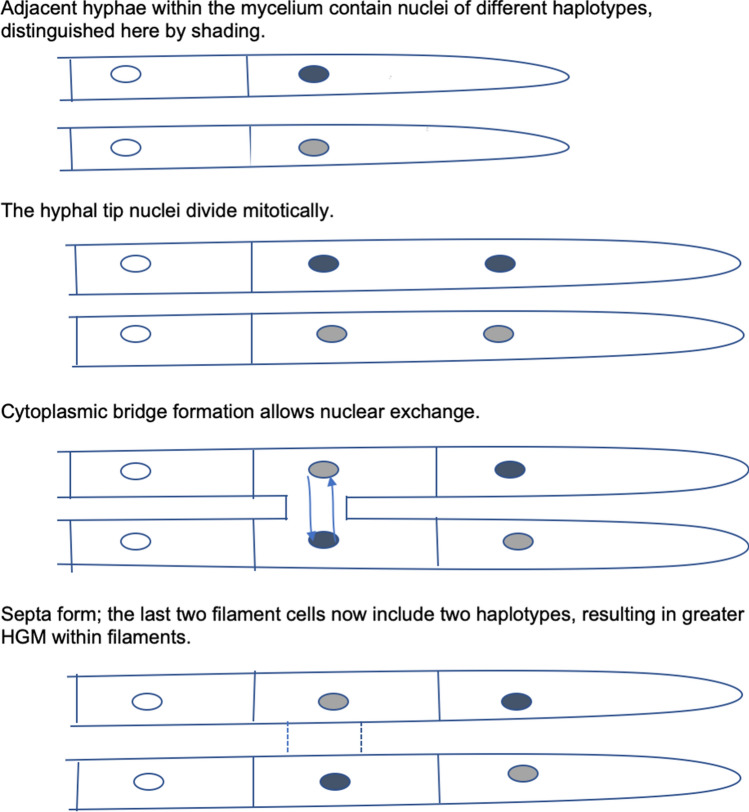
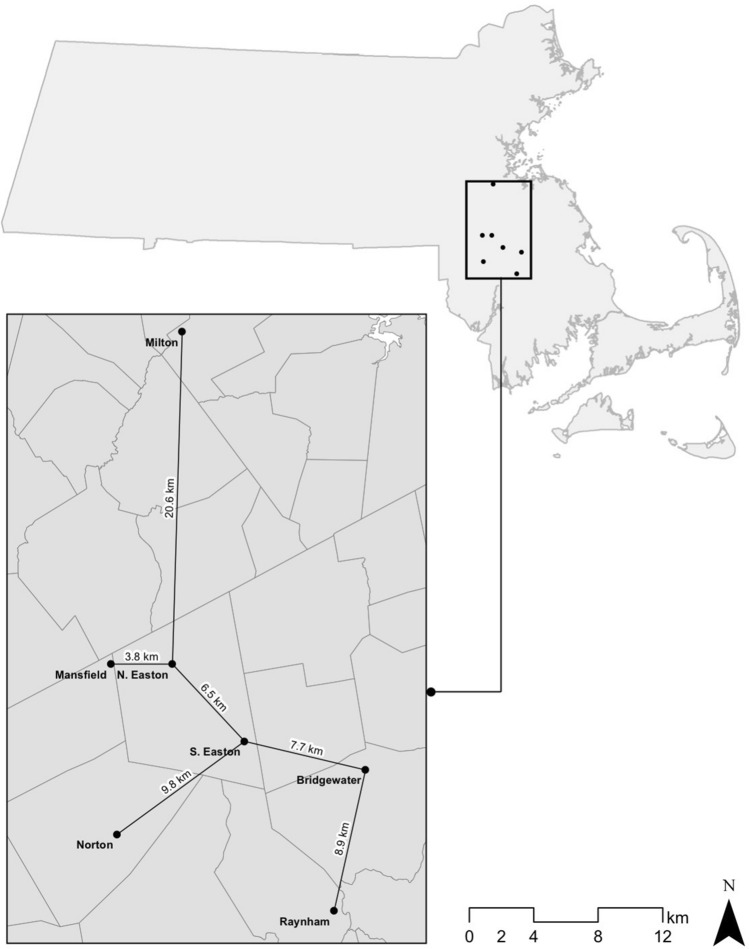
Although cells of mushroom-producing fungi typically contain paired haploid nuclei (n + n), most Armillaria gallica vegetative cells are uninucleate. As vegetative nuclei are produced by fusions of paired haploid nuclei, they are thought to be diploid (2n). Here we report finding haploid vegetative nuclei in A. gallica at multiple sites in southeastern Massachusetts, USA. Sequencing multiple clones of a single-copy gene isolated from single hyphal filaments revealed nuclear heterogeneity both among and within hyphae. Cytoplasmic bridges connected hyphae in field-collected and cultured samples, and we propose nuclear migration through bridges maintains this nuclear heterogeneity. Growth studies demonstrate among- and within-hypha phenotypic variation for growth in response to gallic acid, a plant-produced antifungal compound. The existence of both genetic and phenotypic variation within vegetative hyphae suggests that fungal individuals have the potential to evolve within a single generation in response to environmental variation over time and space.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Raper JR. Genetics of Sexuality in Higher Fungi. New York: The Ronald Press Company; 1966.
-
- Korhonen K. The origin of clamped and clampless basidia in Armillariella ostoyae. Karstenia. 1980;20:23–27. doi: 10.29203/ka.1980.193. - DOI
-
- Grillo R, Korhonen K, Tirro A. The origin of dikaryotic hyphae in fruit bodies of Armillaria borealis and A. tabescens developed in pure culture. In: Johansson M, Stenlid J, editors. Proceedings 8th International Conference on Root and Butt Rots. Uppsala: Swedish University of Agricultural Sciences; 1994. pp. 504–511.
Publication types
MeSH terms
Supplementary concepts
LinkOut - more resources
Full Text Sources
Research Materials
