

Lipid Droplets Maintain Energy Homeostasis and Glioblastoma Growth via Autophagic Release of Stored Fatty Acids
- PMID: 33083736
- PMCID: PMC7549116
- DOI: 10.1016/j.isci.2020.101569
Lipid Droplets Maintain Energy Homeostasis and Glioblastoma Growth via Autophagic Release of Stored Fatty Acids
Abstract
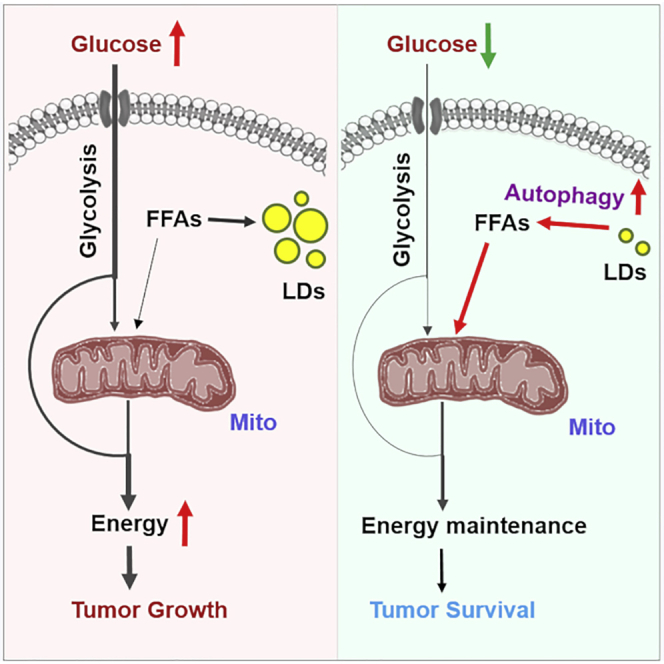
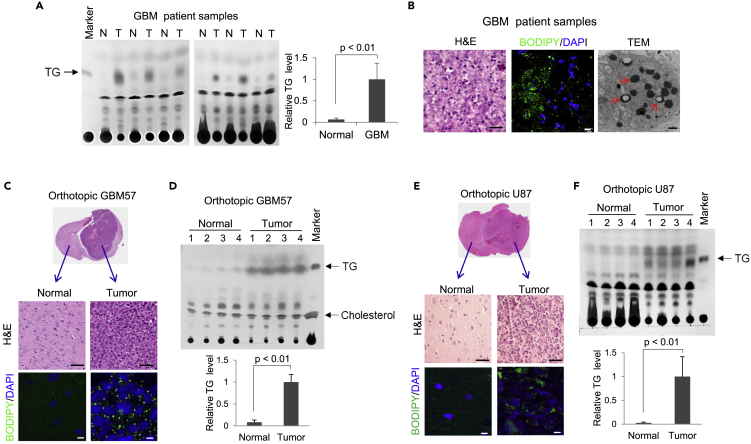
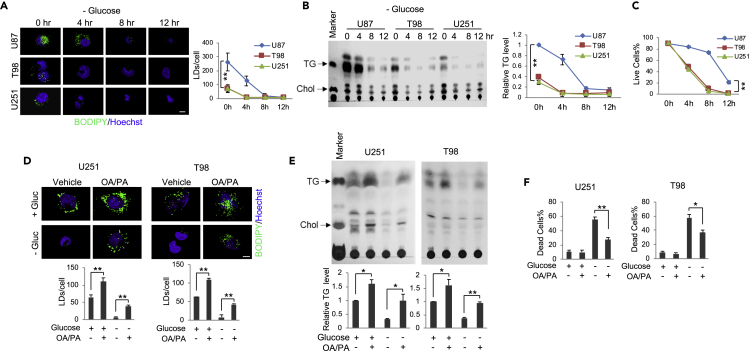
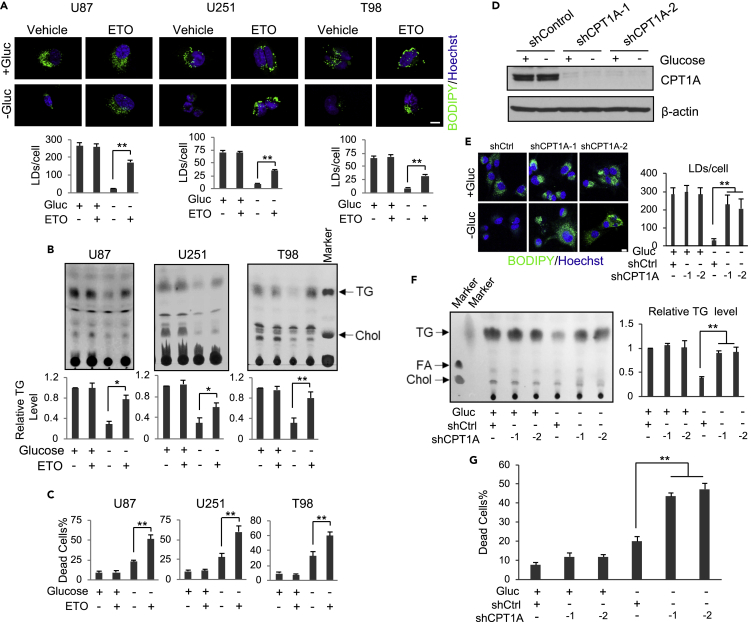
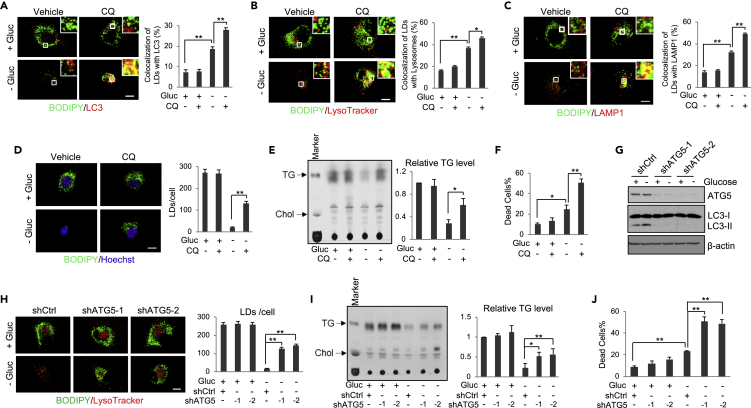
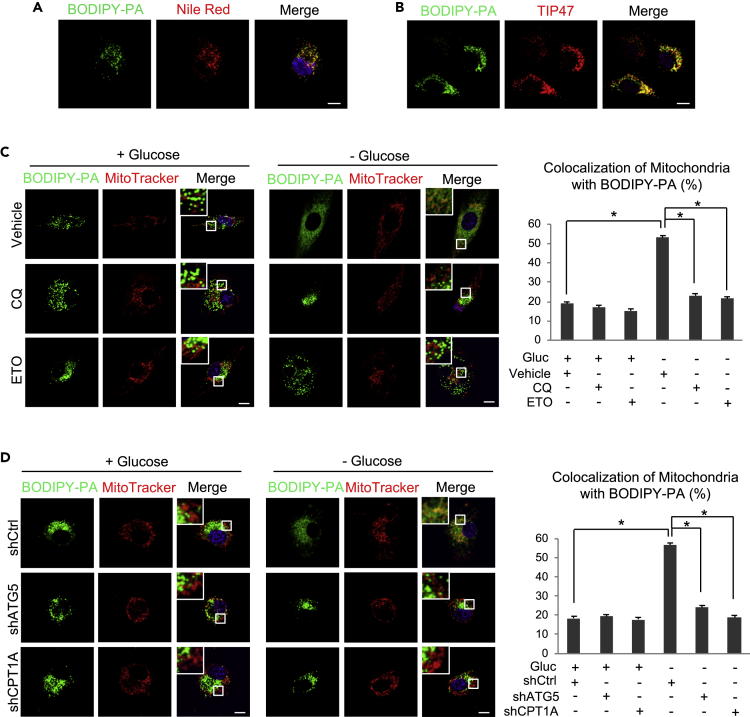
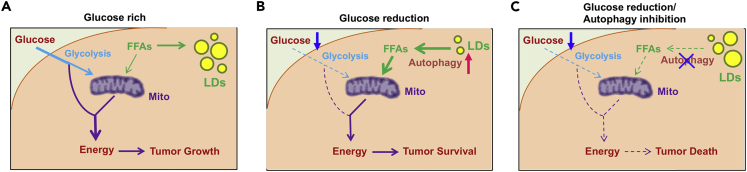
Recently, lipid metabolism reprogramming has been further evidenced in malignancies via the observation of large amounts of lipid droplets (LDs) in human tumors, including in glioblastoma (GBM), the most lethal primary brain tumor. However, the role played by LDs in tumor cells remains unknown. Here, we show that triglycerides (TG), the major components of LDs, serve as a critical energy reservoir to support GBM cell survival. TG/LDs rapidly diminished in GBM cells upon glucose reduction, whereas inhibiting fatty acid oxidation or autophagy resulted in the accumulation of TG/LDs and strongly potentiated GBM cell death. Immunofluorescence imaging and time-lapse videos showed that LDs are hydrolyzed by autophagy to release free fatty acids that mobilize into mitochondria for energy production. Our study demonstrates that autophagy-mediated hydrolysis of TG/LDs maintains energy homeostasis and GBM survival upon glucose reduction, suggesting that limiting TG/LDs utilization might be necessary upon treating GBM.
Keywords: Cancer; Cell Biology; Cellular Physiology.
© 2020 The Author(s).
Conflict of interest statement
The authors declare that there are no conflicts of interest for this manuscript.
Figures
References
-
- Accioly M.T., Pacheco P., Maya-Monteiro C.M., Carrossini N., Robbs B.K., Oliveira S.S., Kaufmann C., Morgado-Diaz J.A., Bozza P.T., Viola J.P. Lipid bodies are reservoirs of cyclooxygenase-2 and sites of prostaglandin-E2 synthesis in colon cancer cells. Cancer Res. 2008;68:1732–1740. - PubMed
-
- Bao S., Wu Q., McLendon R.E., Hao Y., Shi Q., Hjelmeland A.B., Dewhirst M.W., Bigner D.D., Rich J.N. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature. 2006;444:756–760. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
