

microRNA-146a controls age-related bone loss
- PMID: 33085187
- PMCID: PMC7681058
- DOI: 10.1111/acel.13244
microRNA-146a controls age-related bone loss
Abstract
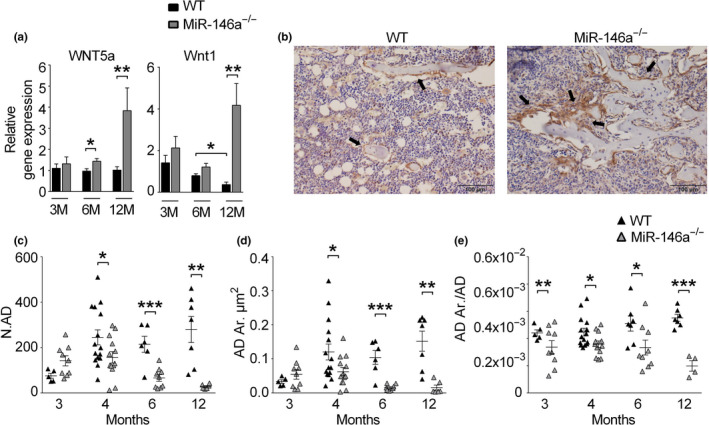
Bone loss is one of the consequences of aging, leading to diseases such as osteoporosis and increased susceptibility to fragility fractures and therefore considerable morbidity and mortality in humans. Here, we identify microRNA-146a (miR-146a) as an essential epigenetic switch controlling bone loss with age. Mice deficient in miR-146a show regular development of their skeleton. However, while WT mice start to lose bone with age, animals deficient in miR-146a continue to accrue bone throughout their life span. Increased bone mass is due to increased generation and activity of osteoblasts in miR-146a-deficient mice as a result of sustained activation of bone anabolic Wnt signaling during aging. Deregulation of the miR-146a target genes Wnt1 and Wnt5a parallels bone accrual and osteoblast generation, which is accompanied by reduced development of bone marrow adiposity. Furthermore, miR-146a-deficient mice are protected from ovariectomy-induced bone loss. In humans, the levels of miR-146a are increased in patients suffering fragility fractures in comparison with those who do not. These data identify miR-146a as a crucial epigenetic temporal regulator which essentially controls bone homeostasis during aging by regulating bone anabolic Wnt signaling. Therefore, miR-146a might be a powerful therapeutic target to prevent age-related bone dysfunctions such as the development of bone marrow adiposity and osteoporosis.
Keywords: aging; bone metabolism; microRNA; osteopetrosis; osteoporosis.
© 2020 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
MH and JG are co‐founders and shareholders of TAmiRNA GmbH, and MH and MW are employed by TAmiRNA GmbH. All other authors have declared that no conflict of interest exists.
Figures




References
-
- Bakker, E. R. M. , Raghoebir, L. , Franken, P. F. , Helvensteijn, W. , van Gurp, L. , Meijlink, F. , van der Valk, M. A. , Rottier, R. J. , Kuipers, E. J. , van Veelen, W. , & Smits, R. (2012). Induced Wnt5a expression perturbs embryonic outgrowth and intestinal elongation, but is well‐tolerated in adult mice. Developmental Biology, 369(1), 91–100. 10.1016/j.ydbio.2012.06.007 - DOI - PubMed
-
- Bernstein, J. , American Academy of Orthopaedic Surgeons , American Academy of Family Physicians , & American Academy of Pediatrics (2003). Musculoskeletal medicine (1st ed.). American Academy of Orthopaedic Surgeons.
-
- Bilkovski, R. , Schulte, D. M. , Oberhauser, F. , Gomolka, M. , Udelhoven, M. , Hettich, M. M. , Roth, B. , Heidenreich, A. , Gutschow, C. , Krone, W. , & Laudes, M. (2010). Role of WNT‐5a in the determination of human mesenchymal stem cells into preadipocytes. Journal of Biological Chemistry, 285(9), 6170–6178. 10.1074/jbc.M109.054338 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
