

Local glutamate-mediated dendritic plateau potentials change the state of the cortical pyramidal neuron
- PMID: 33085562
- PMCID: PMC8087381
- DOI: 10.1152/jn.00734.2019
Local glutamate-mediated dendritic plateau potentials change the state of the cortical pyramidal neuron
Abstract
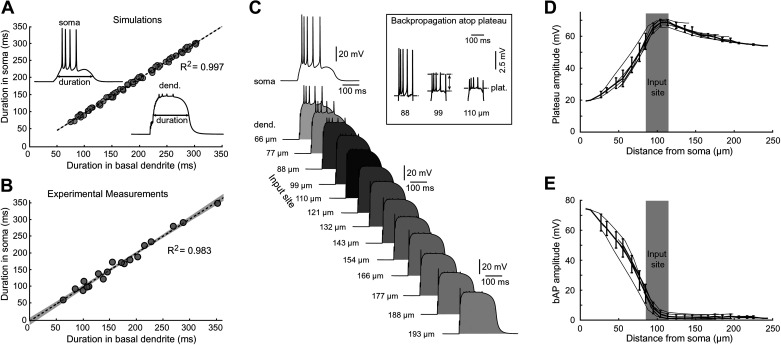
Dendritic spikes in thin dendritic branches (basal and oblique dendrites) are traditionally inferred from spikelets measured in the cell body. Here, we used laser-spot voltage-sensitive dye imaging in cortical pyramidal neurons (rat brain slices) to investigate the voltage waveforms of dendritic potentials occurring in response to spatially restricted glutamatergic inputs. Local dendritic potentials lasted 200-500 ms and propagated to the cell body, where they caused sustained 10- to 20-mV depolarizations. Plateau potentials propagating from dendrite to soma and action potentials propagating from soma to dendrite created complex voltage waveforms in the middle of the thin basal dendrite, comprised of local sodium spikelets, local plateau potentials, and backpropagating action potentials, superimposed on each other. Our model replicated these voltage waveforms across a gradient of glutamatergic stimulation intensities. The model then predicted that somatic input resistance (Rin) and membrane time constant (tau) may be reduced during dendritic plateau potential. We then tested these model predictions in real neurons and found that the model correctly predicted the direction of Rin and tau change but not the magnitude. In summary, dendritic plateau potentials occurring in basal and oblique branches put pyramidal neurons into an activated neuronal state ("prepared state"), characterized by depolarized membrane potential and smaller but faster membrane responses. The prepared state provides a time window of 200-500 ms, during which cortical neurons are particularly excitable and capable of following afferent inputs. At the network level, this predicts that sets of cells with simultaneous plateaus would provide cellular substrate for the formation of functional neuronal ensembles.NEW & NOTEWORTHY In cortical pyramidal neurons, we recorded glutamate-mediated dendritic plateau potentials with voltage imaging and created a computer model that recreated experimental measures from dendrite and cell body. Our model made new predictions, which were then tested in experiments. Plateau potentials profoundly change neuronal state: a plateau potential triggered in one basal dendrite depolarizes the soma and shortens membrane time constant, making the cell more susceptible to firing triggered by other afferent inputs.
Keywords: NMDA spike; ensembles; integration; membrane time constant; nonlinear.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures









References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases