

Innate-adaptive immunity interplay and redox regulation in immune response
- PMID: 33086106
- PMCID: PMC7575795
- DOI: 10.1016/j.redox.2020.101759
Innate-adaptive immunity interplay and redox regulation in immune response
Abstract
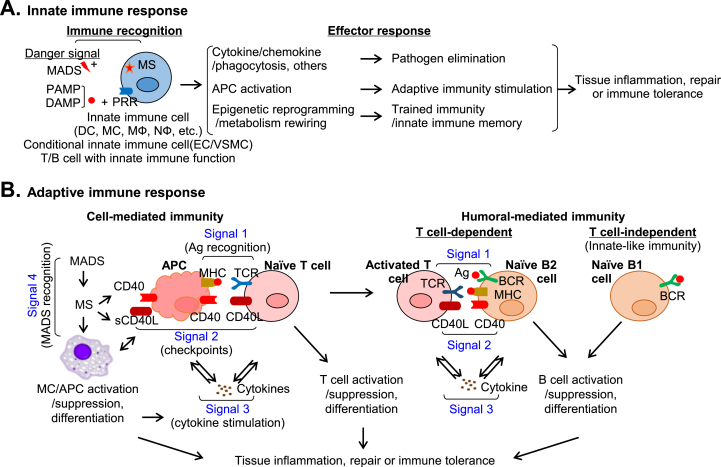
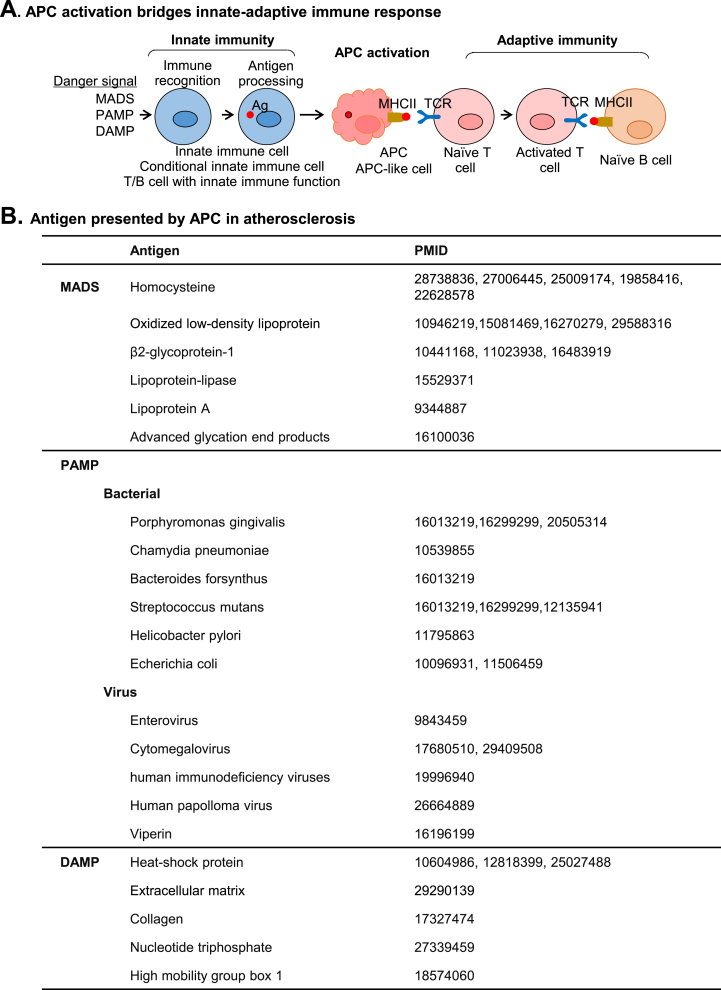
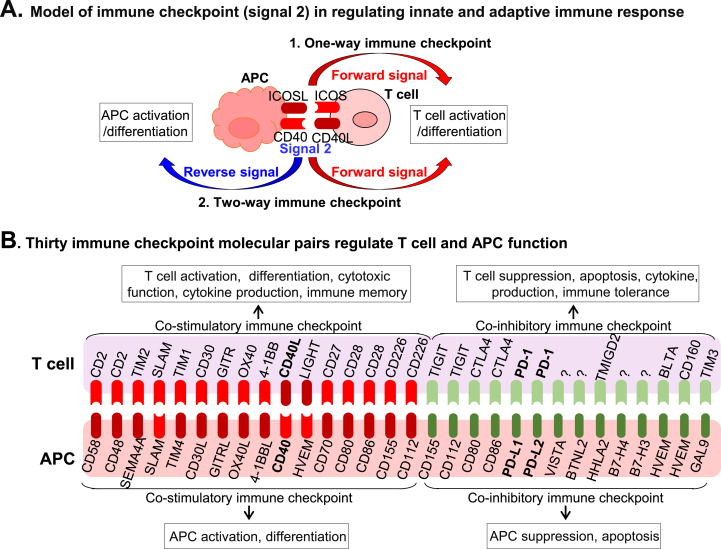
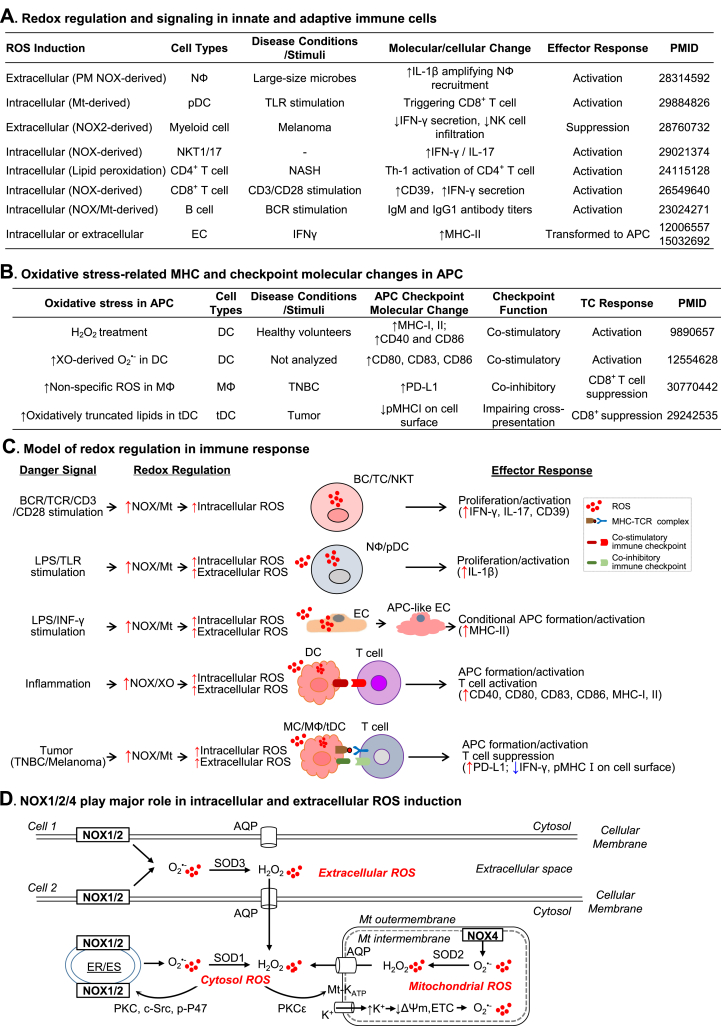
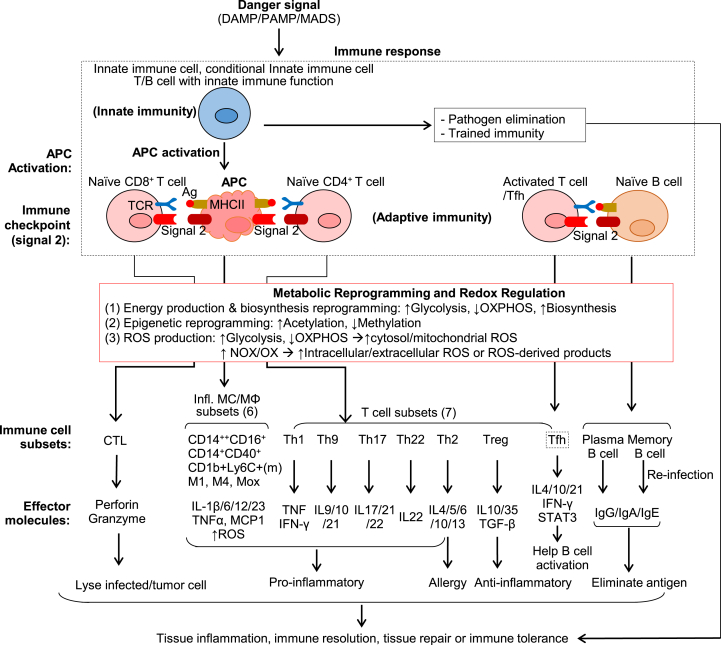
Innate and adaptive immune cell activation and infiltration is the key characteristic of tissue inflammation. The innate immune system is the front line of host defense in which innate immune cells are activated by danger signals, including pathogen- and danger-associated molecular pattern, and metabolite-associated danger signal. Innate immunity activation can directly contribute to tissue inflammation or immune resolution by phagocytosis and secretion of biologically active molecules, or indirectly via antigen-presenting cell (APC) activation-mediated adaptive immune responses. This review article describes the cellular and molecular interplay of innate-adaptive immune systems. Three major mechanisms are emphasized in this article for their role in facilitating innate-adaptive immunity interplay. 1) APC can be formed from classical and conditional innate immune cells to bridge innate-adaptive immune response. 2) Immune checkpoint molecular pairs connect innate and adaptive immune cells to direct one-way and two-way immune checkpoint reactions. 3) Metabolic reprogramming during immune responses leads to excessive cytosolic and mitochondrial reactive oxygen species (ROS) production. Increased NADPH oxidase-derived extracellular and intracellular ROS are mostly responsible for oxidative stress, which contributes to functional changes in immune cells. Further understanding of innate-adaptive immunity interplay and its underlying molecular basis would lead to the identification of therapeutic targets for immunological and inflammatory disease.
Keywords: Adaptive immunity; Immune checkpoint; Immune interplay; Innate immunity; Reactive oxygen species.
Copyright © 2020 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
None.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
