

Integrated proteomics reveals brain-based cerebrospinal fluid biomarkers in asymptomatic and symptomatic Alzheimer's disease
- PMID: 33087358
- PMCID: PMC7577712
- DOI: 10.1126/sciadv.aaz9360
Integrated proteomics reveals brain-based cerebrospinal fluid biomarkers in asymptomatic and symptomatic Alzheimer's disease
Abstract
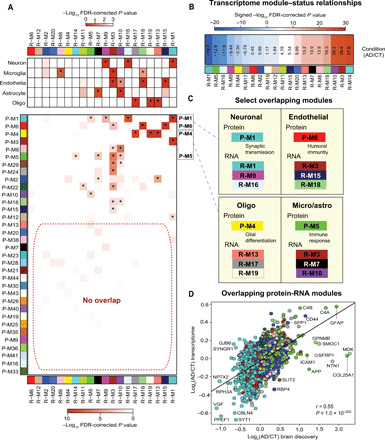
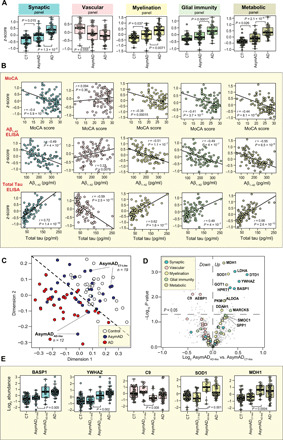
Alzheimer's disease (AD) lacks protein biomarkers reflective of its diverse underlying pathophysiology, hindering diagnostic and therapeutic advancements. Here, we used integrative proteomics to identify cerebrospinal fluid (CSF) biomarkers representing a wide spectrum of AD pathophysiology. Multiplex mass spectrometry identified ~3500 and ~12,000 proteins in AD CSF and brain, respectively. Network analysis of the brain proteome resolved 44 biologically diverse modules, 15 of which overlapped with the CSF proteome. CSF AD markers in these overlapping modules were collapsed into five protein panels representing distinct pathophysiological processes. Synaptic and metabolic panels were decreased in AD brain but increased in CSF, while glial-enriched myelination and immunity panels were increased in brain and CSF. The consistency and disease specificity of panel changes were confirmed in >500 additional CSF samples. These panels also identified biological subpopulations within asymptomatic AD. Overall, these results are a promising step toward a network-based biomarker tool for AD clinical applications.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures




References
-
- Lista S., Zetterberg H., O’Bryant S. E., Blennow K., Hampel H., Evolving relevance of neuroproteomics in Alzheimer’s disease. Methods Mol. Biol. 1598, 101–115 (2017). - PubMed
-
- Castrillo J. I., Lista S., Hampel H., Ritchie C. W., Systems biology methods for Alzheimer’s disease research toward molecular signatures, subtypes, and stages and precision medicine: Application in cohort studies and trials. Methods Mol. Biol. 1750, 31–66 (2018). - PubMed
-
- De Strooper B., Karran E., The cellular phase of Alzheimer’s disease. Cell 164, 603–615 (2016). - PubMed
-
- Blennow K., Hampel H., Weiner M., Zetterberg H., Cerebrospinal fluid and plasma biomarkers in Alzheimer disease. Nat. Rev. Neurol. 6, 131–144 (2010). - PubMed
-
- Jack C. R. Jr., Bennett D. A., Blennow K., Carrillo M. C., Dunn B., Haeberlein S. B., Holtzman D. M., Jagust W., Jessen F., Karlawish J., Liu E., Molinuevo J. L., Montine T., Phelps C., Rankin K. P., Rowe C. C., Scheltens P., Siemers E., Snyder H. M., Sperling R., NIA-AA research framework: Toward a biological definition of Alzheimer’s disease. Alzheimers Dement. 14, 535–562 (2018). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
