

Structured RNA Contaminants in Bacterial Ribo-Seq
- PMID: 33087519
- PMCID: PMC7580957
- DOI: 10.1128/mSphere.00855-20
Structured RNA Contaminants in Bacterial Ribo-Seq
Abstract
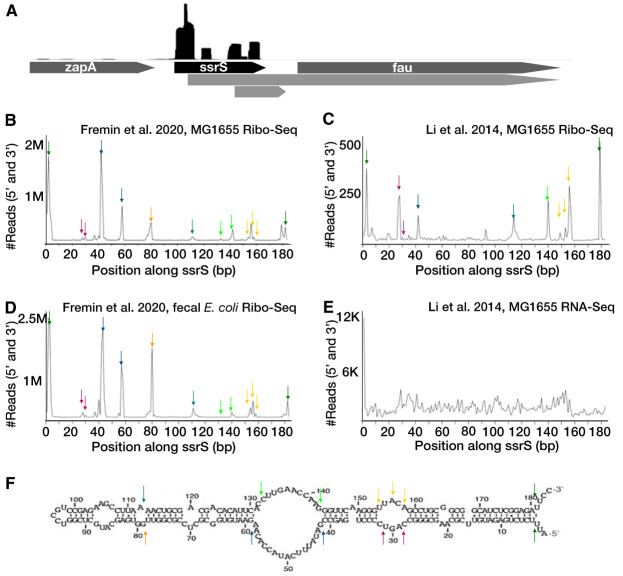
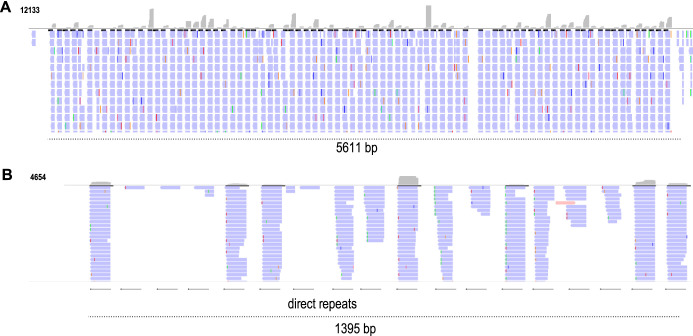
Ribosome profiling (Ribo-Seq) is a powerful method to study translation in bacteria. However, Ribo-Seq signal can be observed across RNAs that one would not expect to be bound by ribosomes. For example, Escherichia coli Ribo-Seq libraries also capture reads from most noncoding RNAs (ncRNAs). While some of these ncRNAs may overlap coding regions, this alone does not explain the majority of observed signal across ncRNAs. These fragments of ncRNAs in Ribo-Seq data pass all size selection steps of the Ribo-Seq protocol and survive hours of micrococcal nuclease (MNase) treatment. In this work, we specifically focus on Ribo-Seq signal across ncRNAs and provide evidence to suggest that RNA structure, as opposed to ribosome binding, protects them from degradation and allows them to persist in the Ribo-Seq sequencing library preparation. By inspecting these "contaminant reads" in bacterial Ribo-Seq, we show that data previously disregarded in bacterial Ribo-Seq experiments may, in fact, be used to gain partial information regarding the in vivo secondary structure of ncRNAs.IMPORTANCE Structured ncRNAs are pivotal mediators of bioregulation in bacteria, and their functions are often reliant on their specific structures. Here, we first inspect Ribo-Seq reads across noncoding regions, identifying contaminant reads in these libraries. We observe that contaminant reads in bacterial Ribo-Seq experiments that are often disregarded, in fact, strongly overlap with structured regions of ncRNAs. We then perform several bioinformatic analyses to determine why these contaminant reads may persist in Ribo-Seq libraries. Finally, we highlight some structured RNA contaminants in Ribo-Seq and support the hypothesis that structures in the RNA protect them from MNase digestion. We conclude that researchers should be cautious when interpreting Ribo-Seq signal as coding without considering signal distribution. These findings also may enable us to partially resolve RNA structures, identify novel structured RNAs, and elucidate RNA structure-function relationships in bacteria at a large scale and in vivo through the reanalysis of existing Ribo-Seq data sets.
Keywords: RNA structure; metagenomics; metatranscriptomics; microbiome.
Copyright © 2020 Fremin and Bhatt.
Figures
Similar articles
-
Recommendations for bacterial ribosome profiling experiments based on bioinformatic evaluation of published data.J Biol Chem. 2020 Jul 3;295(27):8999-9011. doi: 10.1074/jbc.RA119.012161. Epub 2020 May 8. J Biol Chem. 2020. PMID: 32385111 Free PMC article.
-
Comparison of RNA-Seq by poly (A) capture, ribosomal RNA depletion, and DNA microarray for expression profiling.BMC Genomics. 2014 Jun 2;15(1):419. doi: 10.1186/1471-2164-15-419. BMC Genomics. 2014. PMID: 24888378 Free PMC article.
-
Understanding the proteome encoded by "non-coding RNAs": new insights into human genome.Sci China Life Sci. 2020 Jul;63(7):986-995. doi: 10.1007/s11427-019-1677-8. Epub 2020 Apr 20. Sci China Life Sci. 2020. PMID: 32318910 Review.
-
Scikit-ribo Enables Accurate Estimation and Robust Modeling of Translation Dynamics at Codon Resolution.Cell Syst. 2018 Feb 28;6(2):180-191.e4. doi: 10.1016/j.cels.2017.12.007. Epub 2018 Jan 17. Cell Syst. 2018. PMID: 29361467 Free PMC article.
-
Ribosomal profiling adds new coding sequences to the proteome.Biochem Soc Trans. 2015 Dec;43(6):1271-6. doi: 10.1042/BST20150170. Biochem Soc Trans. 2015. PMID: 26614672 Review.
Cited by
-
Exposing the small protein load of bacterial life.FEMS Microbiol Rev. 2023 Nov 1;47(6):fuad063. doi: 10.1093/femsre/fuad063. FEMS Microbiol Rev. 2023. PMID: 38012116 Free PMC article. Review.
-
Comparative genomics identifies thousands of candidate structured RNAs in human microbiomes.Genome Biol. 2021 Apr 12;22(1):100. doi: 10.1186/s13059-021-02319-w. Genome Biol. 2021. PMID: 33845850 Free PMC article.
-
High-Resolution Small RNAs Landscape Provides Insights into Alkane Adaptation in the Marine Alkane-Degrader Alcanivorax dieselolei B-5.Int J Mol Sci. 2022 Dec 15;23(24):15995. doi: 10.3390/ijms232415995. Int J Mol Sci. 2022. PMID: 36555635 Free PMC article.
-
What can Ribo-seq and proteomics tell us about the non-canonical proteome?bioRxiv [Preprint]. 2023 May 18:2023.05.16.541049. doi: 10.1101/2023.05.16.541049. bioRxiv. 2023. Update in: Mol Cell Proteomics. 2023 Sep;22(9):100631. doi: 10.1016/j.mcpro.2023.100631. PMID: 37292611 Free PMC article. Updated. Preprint.
-
RNA-seq and Ribosome Profiling Reveal the Translational Landscape of Rice in Response to Rice Stripe Virus Infection.Viruses. 2024 Nov 29;16(12):1866. doi: 10.3390/v16121866. Viruses. 2024. PMID: 39772176 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
