

Cell morphology maintenance in Bacillus subtilis through balanced peptidoglycan synthesis and hydrolysis
- PMID: 33087775
- PMCID: PMC7578834
- DOI: 10.1038/s41598-020-74609-5
Cell morphology maintenance in Bacillus subtilis through balanced peptidoglycan synthesis and hydrolysis
Erratum in
-
Publisher Correction: Cell morphology maintenance in Bacillus subtilis through balanced peptidoglycan synthesis and hydrolysis.Sci Rep. 2021 Mar 11;11(1):6156. doi: 10.1038/s41598-021-85465-2. Sci Rep. 2021. PMID: 33707645 Free PMC article. No abstract available.
Abstract
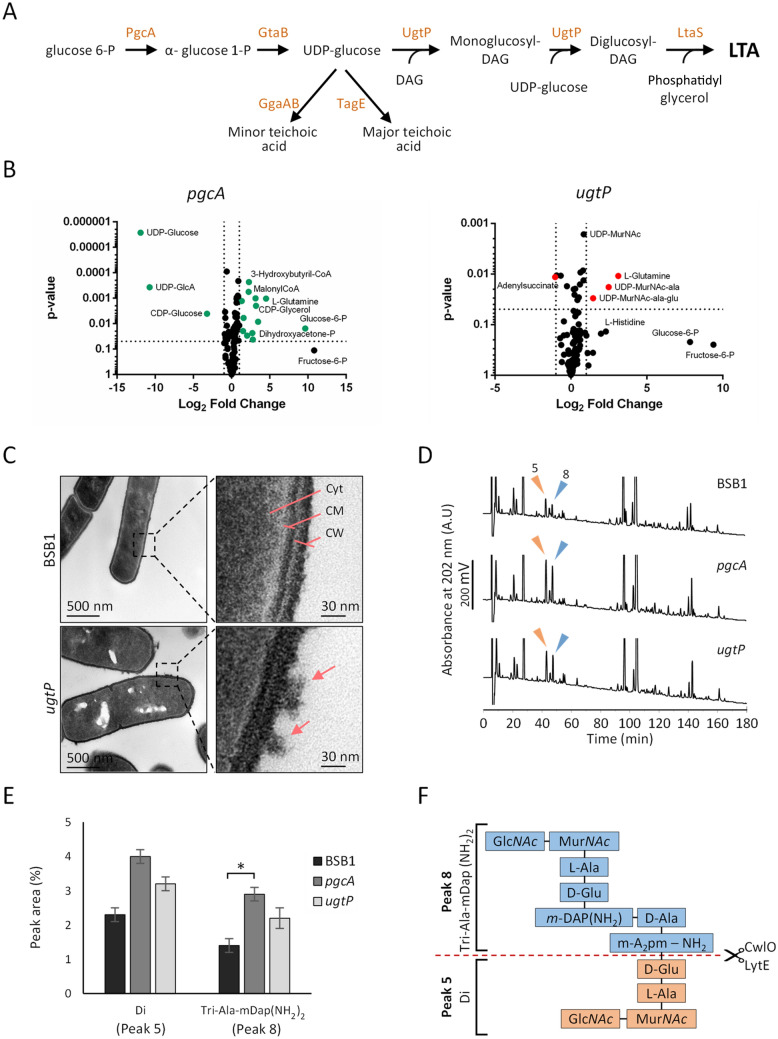
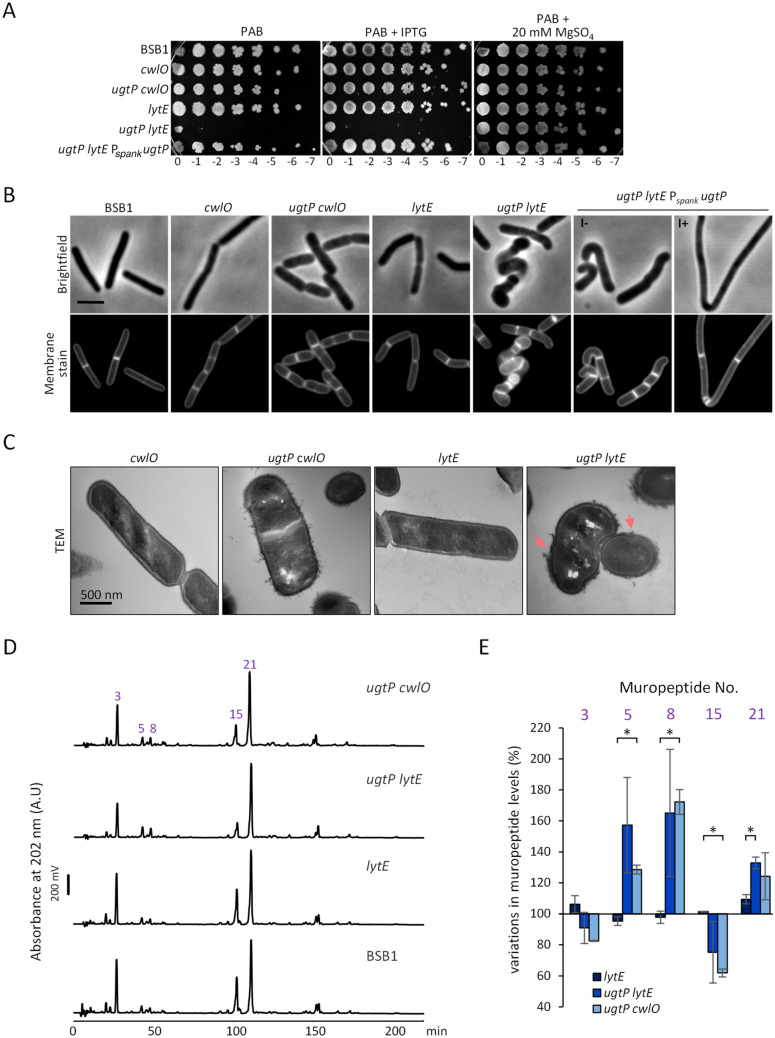
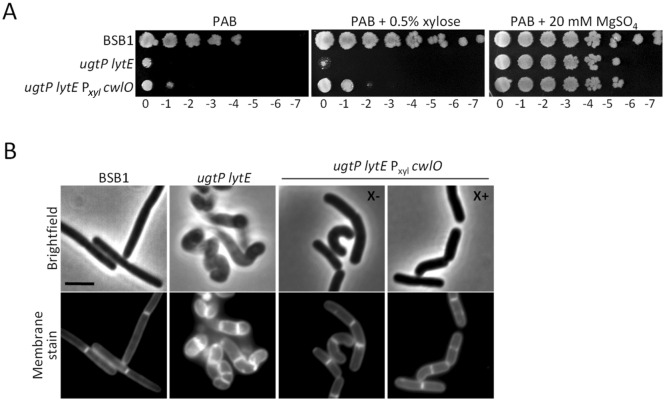
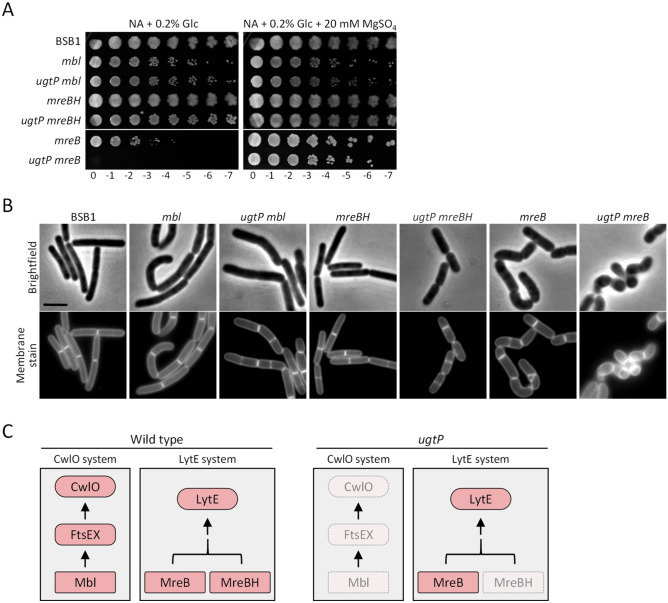
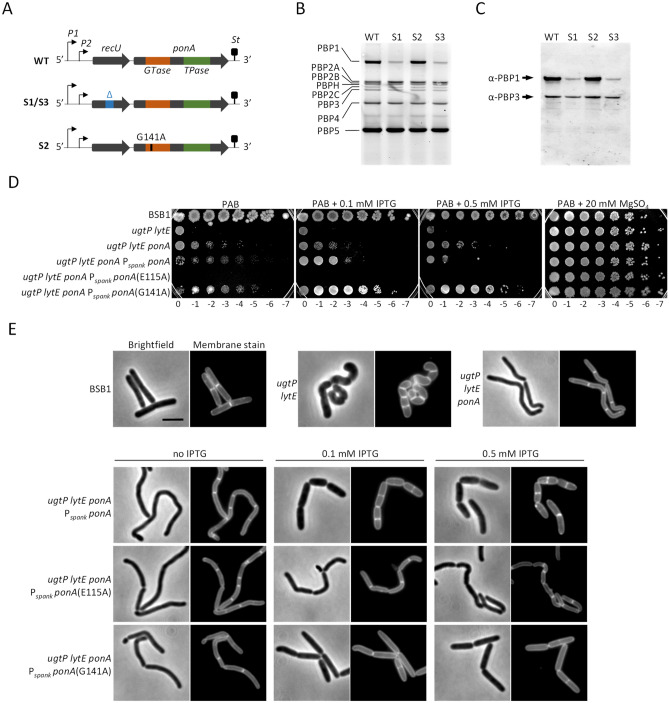
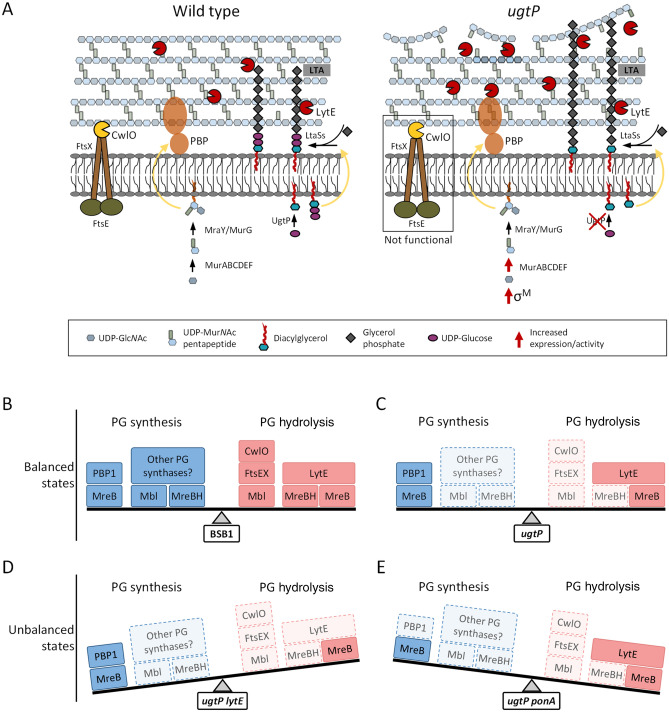
The peptidoglycan layer is responsible for maintaining bacterial cell shape and permitting cell division. Cell wall growth is facilitated by peptidoglycan synthases and hydrolases and is potentially modulated by components of the central carbon metabolism. In Bacillus subtilis, UgtP synthesises the glucolipid precursor for lipoteichoic acid and has been suggested to function as a metabolic sensor governing cell size. Here we show that ugtP mutant cells have increased levels of cell wall precursors and changes in their peptidoglycan that suggest elevated DL-endopeptidase activity. The additional deletion of lytE, encoding a DL-endopeptidase important for cell elongation, in the ugtP mutant background produced cells with severe shape defects. Interestingly, the ugtP lytE mutant recovered normal rod-shape by acquiring mutations that decreased the expression of the peptidoglycan synthase PBP1. Together our results suggest that cells lacking ugtP must re-adjust the balance between peptidoglycan synthesis and hydrolysis to maintain proper cell morphology.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Teichoic Acid Polymers Affect Expression and Localization of dl-Endopeptidase LytE Required for Lateral Cell Wall Hydrolysis in Bacillus subtilis.J Bacteriol. 2016 May 13;198(11):1585-1594. doi: 10.1128/JB.00003-16. Print 2016 Jun 1. J Bacteriol. 2016. PMID: 27002131 Free PMC article.
-
Digestion of peptidoglycan near the cross-link is necessary for the growth of Bacillus subtilis.Microbiology (Reading). 2018 Mar;164(3):299-307. doi: 10.1099/mic.0.000614. Epub 2018 Jan 25. Microbiology (Reading). 2018. PMID: 29458657
-
The WalR-WalK Signaling Pathway Modulates the Activities of both CwlO and LytE through Control of the Peptidoglycan Deacetylase PdaC in Bacillus subtilis.J Bacteriol. 2022 Feb 15;204(2):e0053321. doi: 10.1128/JB.00533-21. Epub 2021 Dec 6. J Bacteriol. 2022. PMID: 34871030 Free PMC article.
-
Shape determination in Bacillus subtilis.Curr Opin Microbiol. 2007 Dec;10(6):611-6. doi: 10.1016/j.mib.2007.09.008. Epub 2007 Nov 5. Curr Opin Microbiol. 2007. PMID: 17981078 Review.
-
From the regulation of peptidoglycan synthesis to bacterial growth and morphology.Nat Rev Microbiol. 2011 Dec 28;10(2):123-36. doi: 10.1038/nrmicro2677. Nat Rev Microbiol. 2011. PMID: 22203377 Free PMC article. Review.
Cited by
-
Bioactivity and genome analysis of Bacillus amyloliquefaciens GL18 isolated from the rhizosphere of Kobresia myosuroides in an alpine meadow.Antonie Van Leeuwenhoek. 2024 Jan 8;117(1):16. doi: 10.1007/s10482-023-01917-x. Antonie Van Leeuwenhoek. 2024. PMID: 38189906
-
Length-based separation of Bacillus subtilis bacterial populations by viscoelastic microfluidics.Microsyst Nanoeng. 2022 Jan 19;8:7. doi: 10.1038/s41378-021-00333-3. eCollection 2022. Microsyst Nanoeng. 2022. PMID: 35127130 Free PMC article.
-
Metabolic Control of Cell Elongation and Cell Division in Bacillus subtilis.Front Microbiol. 2021 Jun 25;12:697930. doi: 10.3389/fmicb.2021.697930. eCollection 2021. Front Microbiol. 2021. PMID: 34248920 Free PMC article. Review.
-
Immunostimulatory activity of inactivated environmental Bacillus isolates and their endospores.Sci Rep. 2025 Aug 20;15(1):30604. doi: 10.1038/s41598-025-12833-7. Sci Rep. 2025. PMID: 40835982 Free PMC article.
-
GpsB interacts with FtsZ in multiple species and may serve as an accessory Z-ring anchor.Mol Biol Cell. 2025 Jan 1;36(1):ar10. doi: 10.1091/mbc.E24-07-0302. Epub 2024 Nov 27. Mol Biol Cell. 2025. PMID: 39602291 Free PMC article.
References
-
- Vollmer W, Blanot D, de Pedro MA. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008;32:149–167. - PubMed
-
- Bhavsar AP, Brown ED. Cell wall assembly in Bacillus subtilis: How spirals and spaces challenge paradigms. Mol. Microbiol. 2006;60:1077–1090. - PubMed
-
- Lovering AL, Safadi SS, Strynadka NCJ. Structural perspective of peptidoglycan biosynthesis and assembly. Annu. Rev. Biochem. 2012;81:451–478. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
