

Structure-Based Design with Tag-Based Purification and In-Process Biotinylation Enable Streamlined Development of SARS-CoV-2 Spike Molecular Probes
- PMID: 33091382
- PMCID: PMC7550166
- DOI: 10.1016/j.celrep.2020.108322
Structure-Based Design with Tag-Based Purification and In-Process Biotinylation Enable Streamlined Development of SARS-CoV-2 Spike Molecular Probes
Abstract
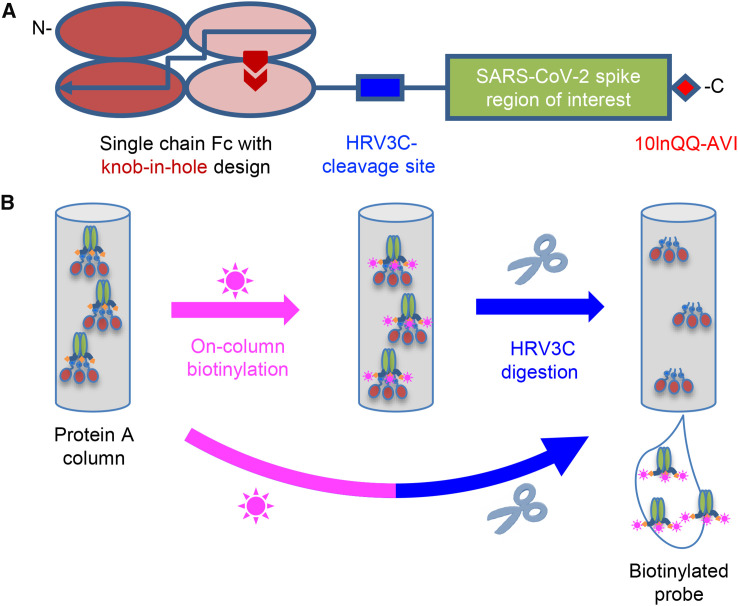
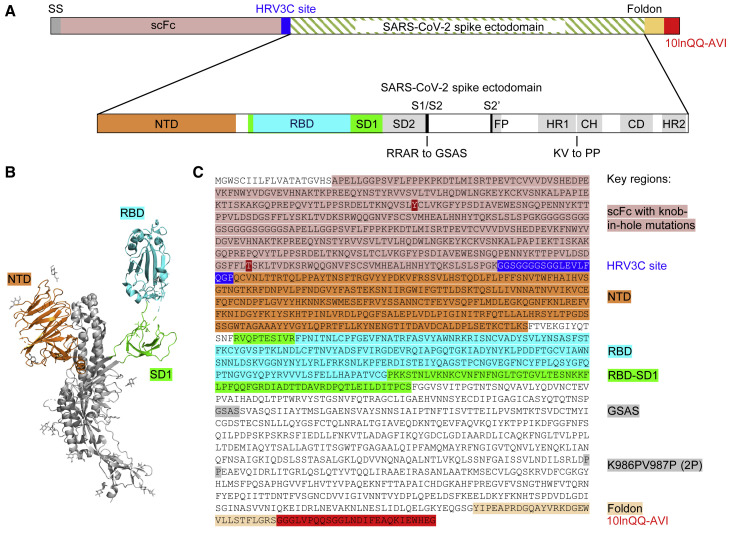
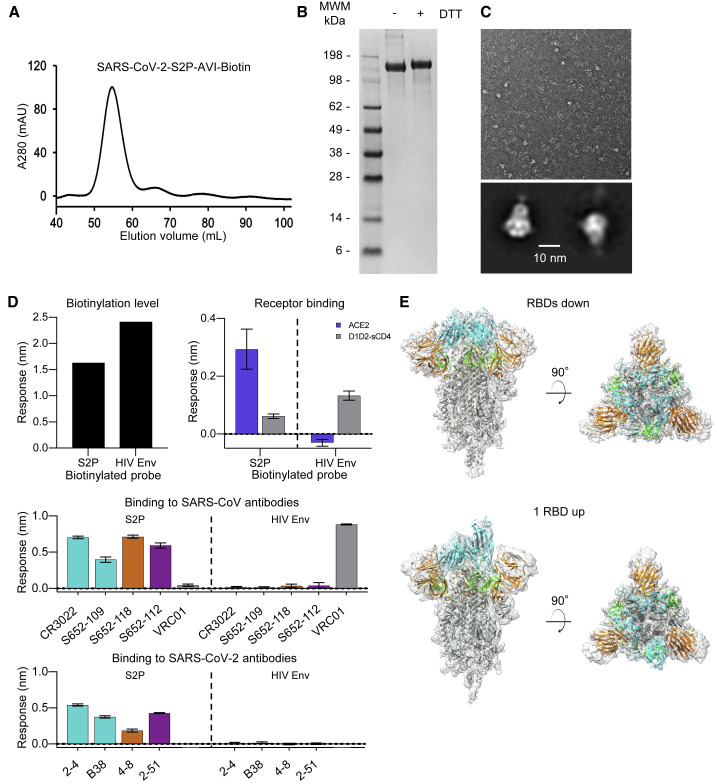
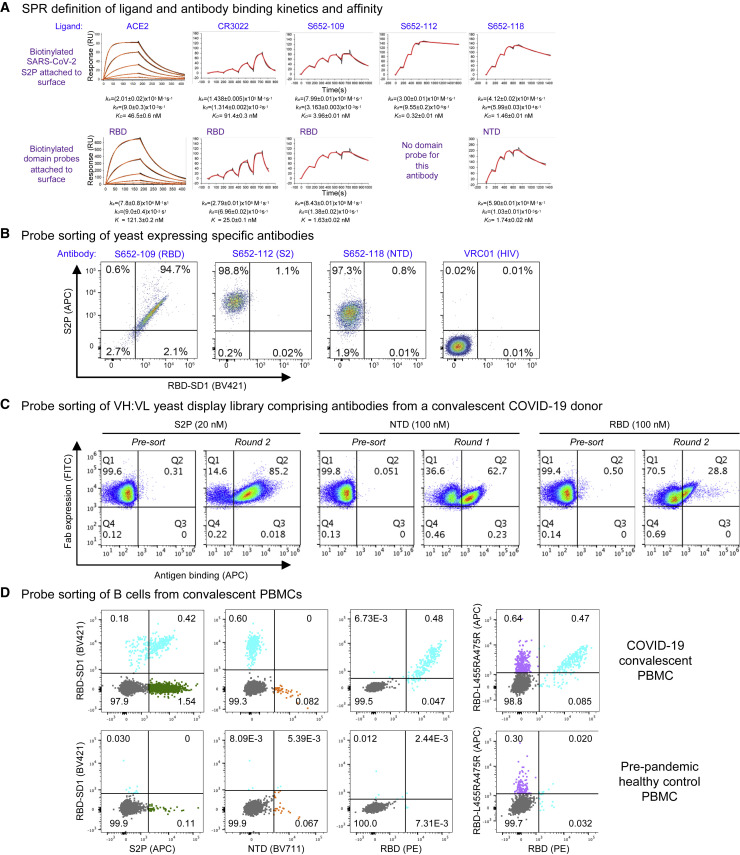
Biotin-labeled molecular probes, comprising specific regions of the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike, would be helpful in the isolation and characterization of antibodies targeting this recently emerged pathogen. Here, we design constructs incorporating an N-terminal purification tag, a site-specific protease-cleavage site, the probe region of interest, and a C-terminal sequence targeted by biotin ligase. Probe regions include full-length spike ectodomain as well as various subregions, and we also design mutants that eliminate recognition of the angiotensin-converting enzyme 2 (ACE2) receptor. Yields of biotin-labeled probes from transient transfection range from ∼0.5 mg/L for the complete ectodomain to >5 mg/L for several subregions. Probes are characterized for antigenicity and ACE2 recognition, and the structure of the spike ectodomain probe is determined by cryoelectron microscopy. We also characterize antibody-binding specificities and cell-sorting capabilities of the biotinylated probes. Altogether, structure-based design coupled to efficient purification and biotinylation processes can thus enable streamlined development of SARS-CoV-2 spike ectodomain probes.
Keywords: COVID-19; HRV3C protease; antibody; biotinylated probe; coronavirus disease 2019; human rhinovirus 3C; single-chain Fc; structure-based design.
Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests The authors declare no competing interest.
Figures



Update of
-
Structure-Based Design with Tag-Based Purification and In-Process Biotinylation Enable Streamlined Development of SARS-CoV-2 Spike Molecular Probes.bioRxiv [Preprint]. 2020 Jun 23:2020.06.22.166033. doi: 10.1101/2020.06.22.166033. bioRxiv. 2020. Update in: Cell Rep. 2020 Oct 27;33(4):108322. doi: 10.1016/j.celrep.2020.108322. PMID: 32596696 Free PMC article. Updated. Preprint.
-
Structure-Based Design with Tag-Based Purification and In-Process Biotinylation Enable Streamlined Development of SARS-CoV-2 Spike Molecular Probes.SSRN [Preprint]. 2020 Jul 21:3639618. doi: 10.2139/ssrn.3639618. SSRN. 2020. Update in: Cell Rep. 2020 Oct 27;33(4):108322. doi: 10.1016/j.celrep.2020.108322. PMID: 32742241 Free PMC article. Updated. Preprint.
References
-
- Adams P.D., Gopal K., Grosse-Kunstleve R.W., Hung L.W., Ioerger T.R., McCoy A.J., Moriarty N.W., Pai R.K., Read R.J., Romo T.D. Recent developments in the PHENIX software for automated crystallographic structure determination. J. Synchrotron Radiat. 2004;11:53–55. - PubMed
-
- Antoniou G., Papakyriacou I., Papaneophytou C. Optimization of Soluble Expression and Purification of Recombinant Human Rhinovirus Type-14 3C Protease Using Statistically Designed Experiments: Isolation and Characterization of the Enzyme. Mol. Biotechnol. 2017;59:407–424. - PubMed
-
- Callaway E. The race for coronavirus vaccines: a graphical guide. Nature. 2020;580:576–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
