

Copper metabolism as a unique vulnerability in cancer
- PMID: 33091507
- PMCID: PMC7779655
- DOI: 10.1016/j.bbamcr.2020.118893
Copper metabolism as a unique vulnerability in cancer
Abstract
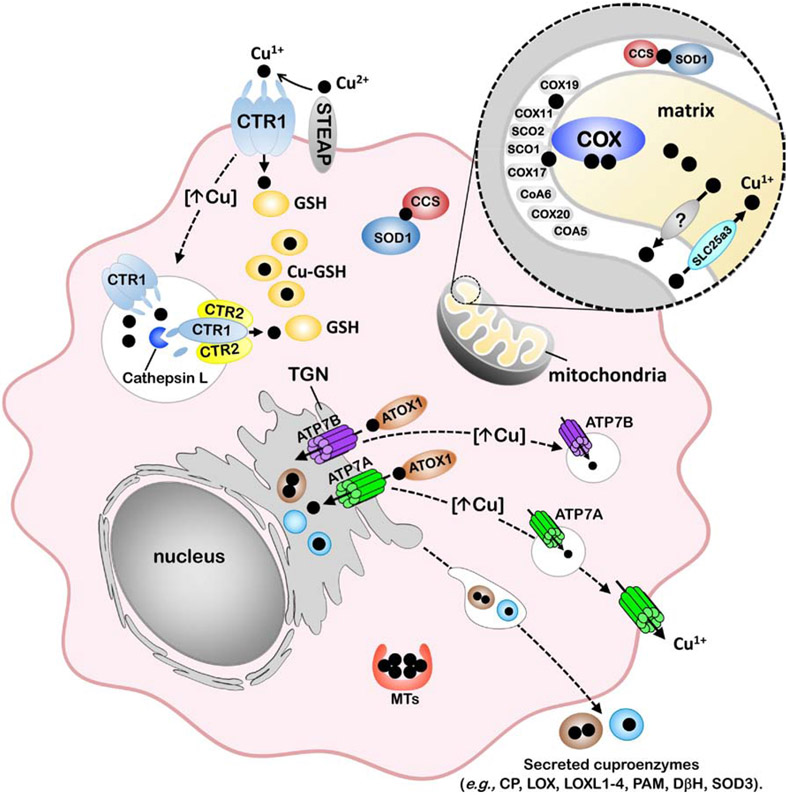
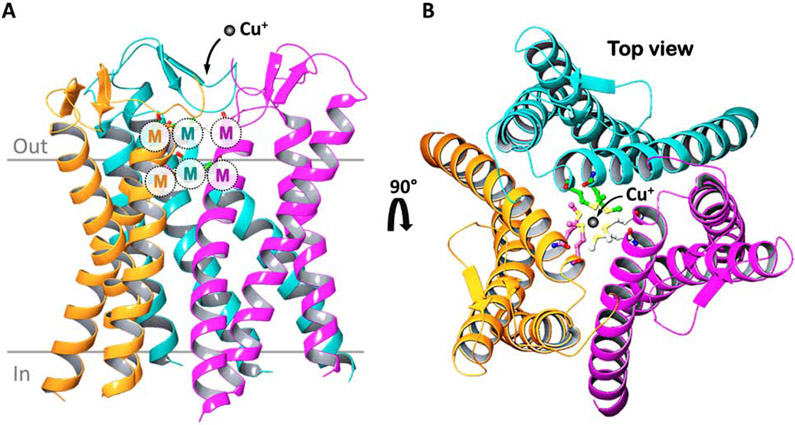
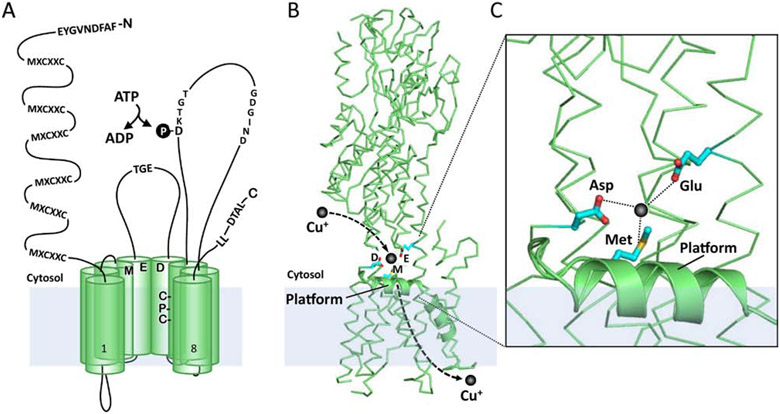
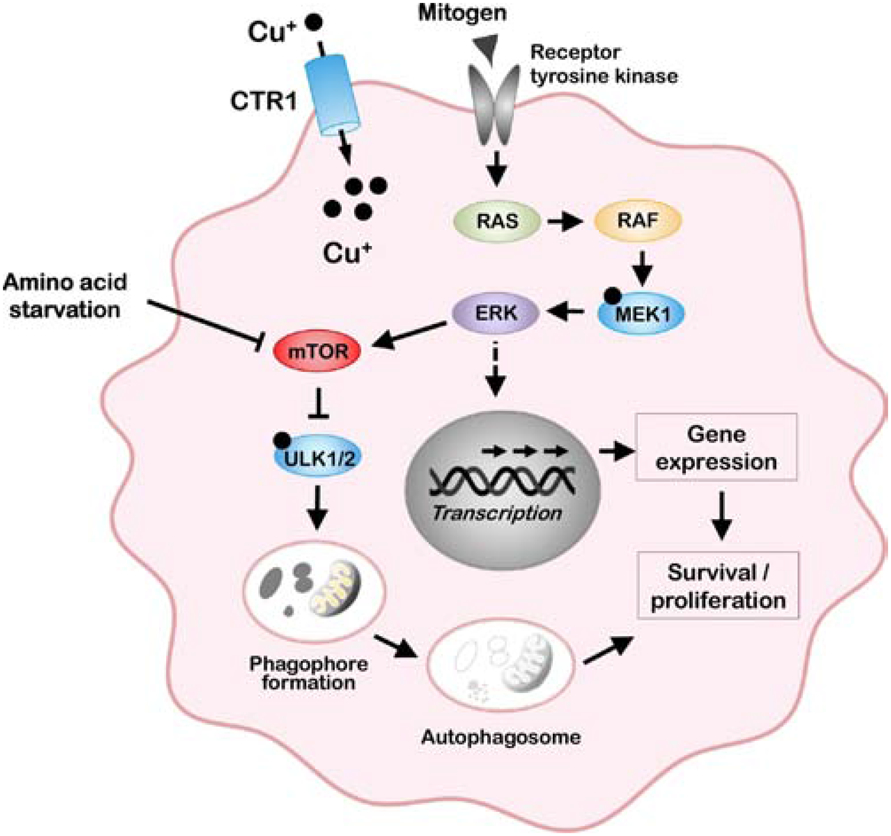
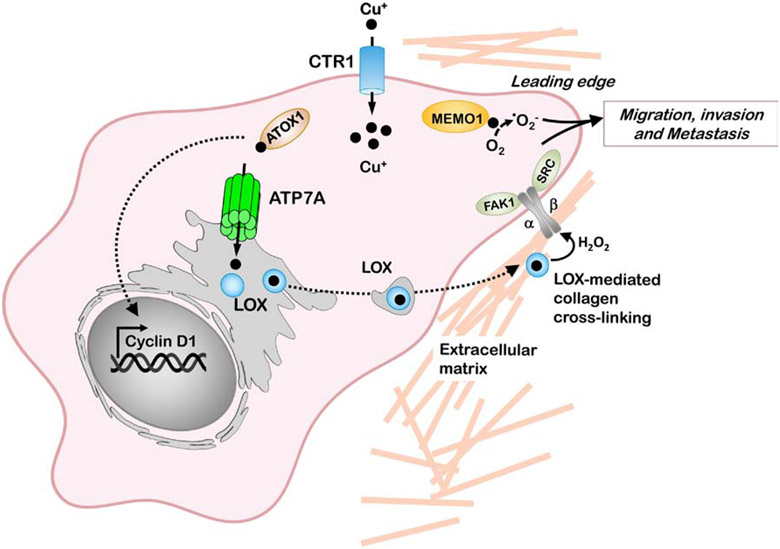
The last 25 years have witnessed tremendous progress in identifying and characterizing proteins that regulate the uptake, intracellular trafficking and export of copper. Although dietary copper is required in trace amounts, sufficient quantities of this metal are needed to sustain growth and development in humans and other mammals. However, copper is also a rate-limiting nutrient for the growth and proliferation of cancer cells. Oral copper chelators taken with food have been shown to confer anti-neoplastic and anti-metastatic benefits in animals and humans. Recent studies have begun to identify specific roles for copper in pathways of oncogenic signaling and resistance to anti-neoplastic drugs. Here, we review the general mechanisms of cellular copper homeostasis and discuss roles of copper in cancer progression, highlighting metabolic vulnerabilities that may be targetable in the development of anticancer therapies.
Keywords: Cancer; Cisplatin chemotherapy; Copper chelation; Copper homeostasis; Metallochaperone; Nutrition.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Ochiai EI, Copper and the biological evolution, Biosystems, 16 (1983) 81–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
