

BRD4 (Bromodomain-Containing Protein 4) Interacts with GATA4 (GATA Binding Protein 4) to Govern Mitochondrial Homeostasis in Adult Cardiomyocytes
- PMID: 33094644
- PMCID: PMC7736290
- DOI: 10.1161/CIRCULATIONAHA.120.047753
BRD4 (Bromodomain-Containing Protein 4) Interacts with GATA4 (GATA Binding Protein 4) to Govern Mitochondrial Homeostasis in Adult Cardiomyocytes
Abstract
Background: Gene regulatory networks control tissue homeostasis and disease progression in a cell type-specific manner. Ubiquitously expressed chromatin regulators modulate these networks, yet the mechanisms governing how tissue specificity of their function is achieved are poorly understood. BRD4 (bromodomain-containing protein 4), a member of the BET (bromo- and extraterminal domain) family of ubiquitously expressed acetyl-lysine reader proteins, plays a pivotal role as a coactivator of enhancer signaling across diverse tissue types in both health and disease and has been implicated as a pharmacological target in heart failure. However, the cell-specific role of BRD4 in adult cardiomyocytes remains unknown.
Methods: We combined conditional mouse genetics, unbiased transcriptomic and epigenomic analyses, and classic molecular biology and biochemical approaches to understand the mechanism by which BRD4 in adult cardiomyocyte homeostasis.
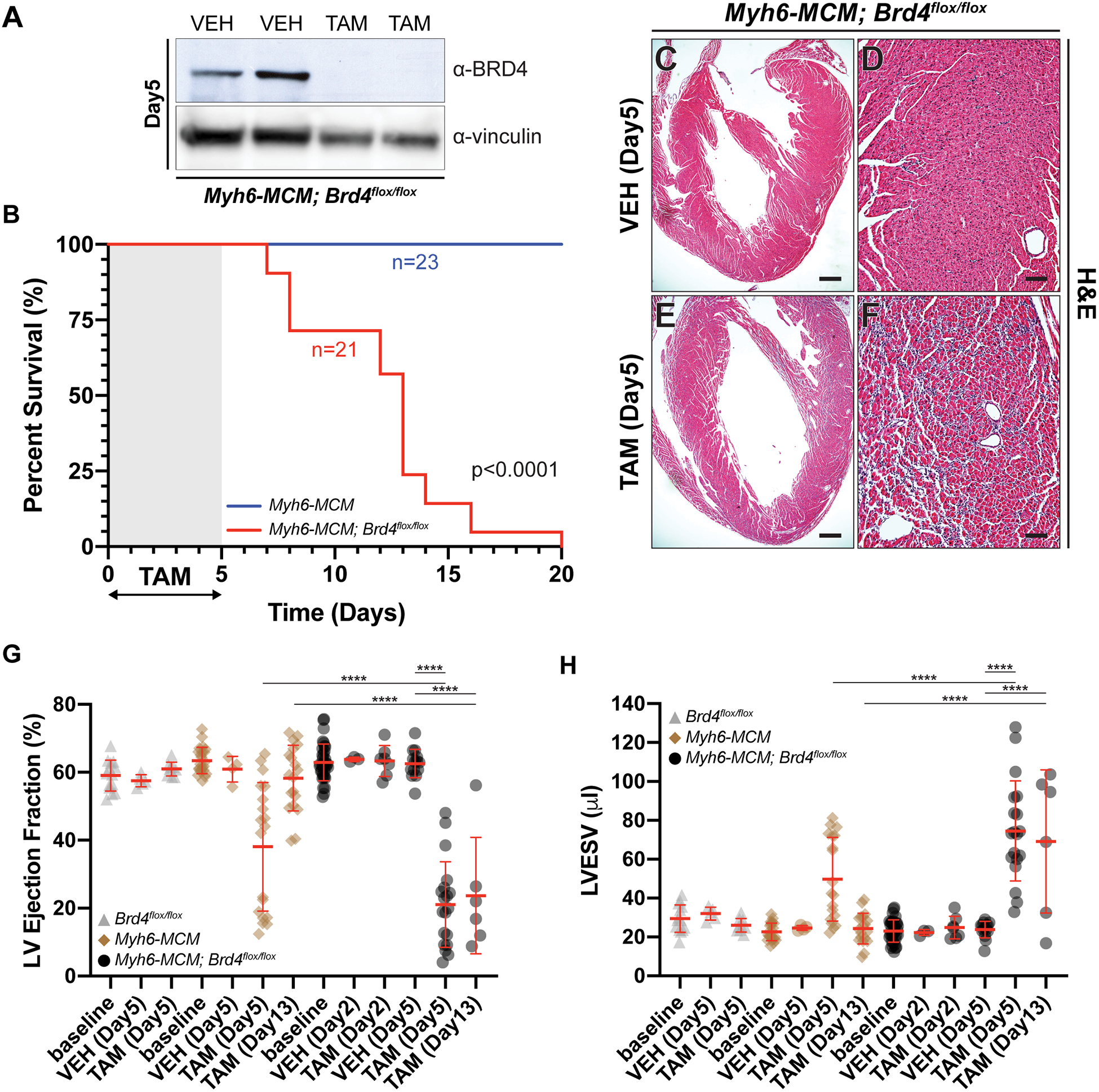
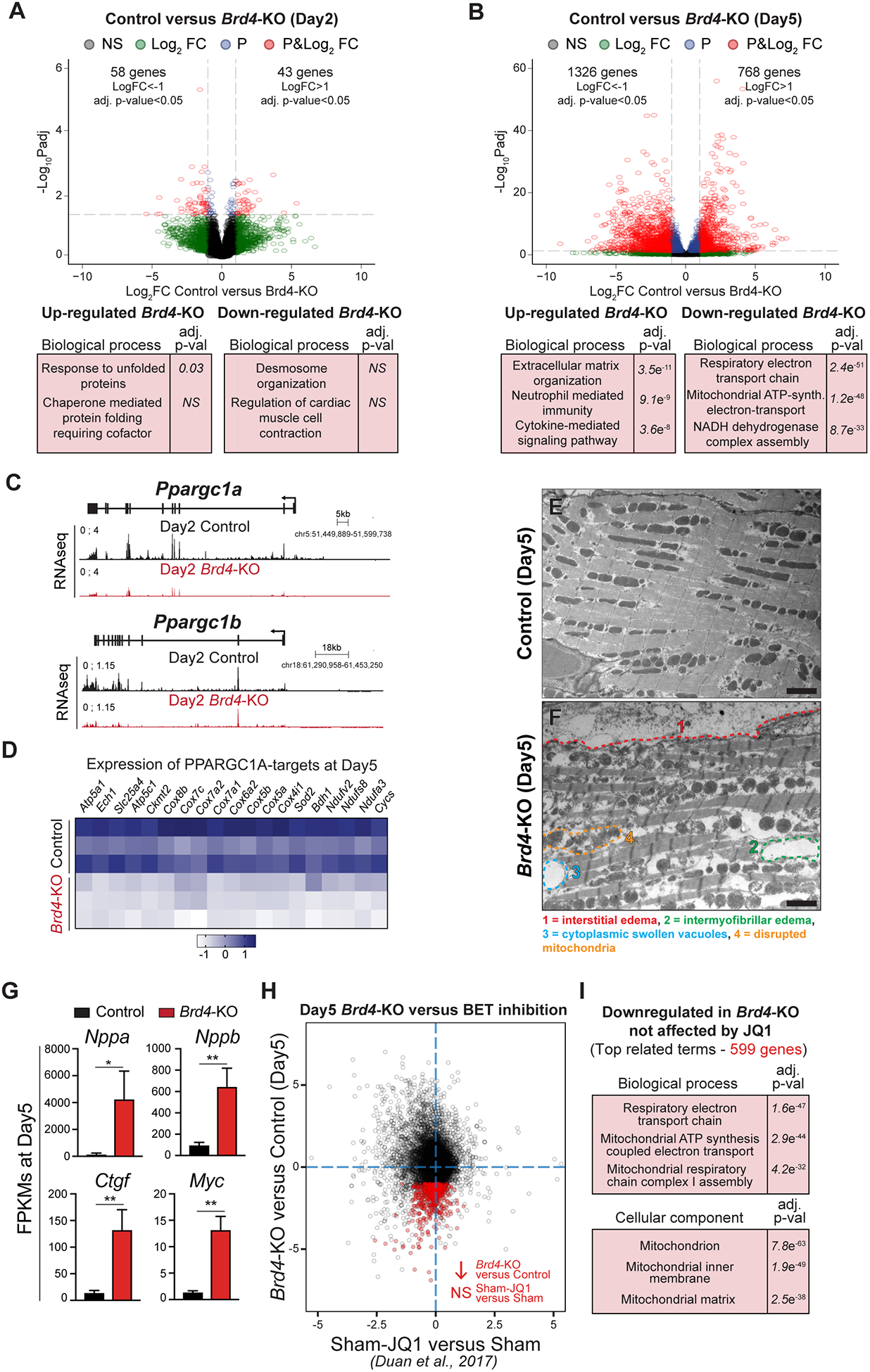
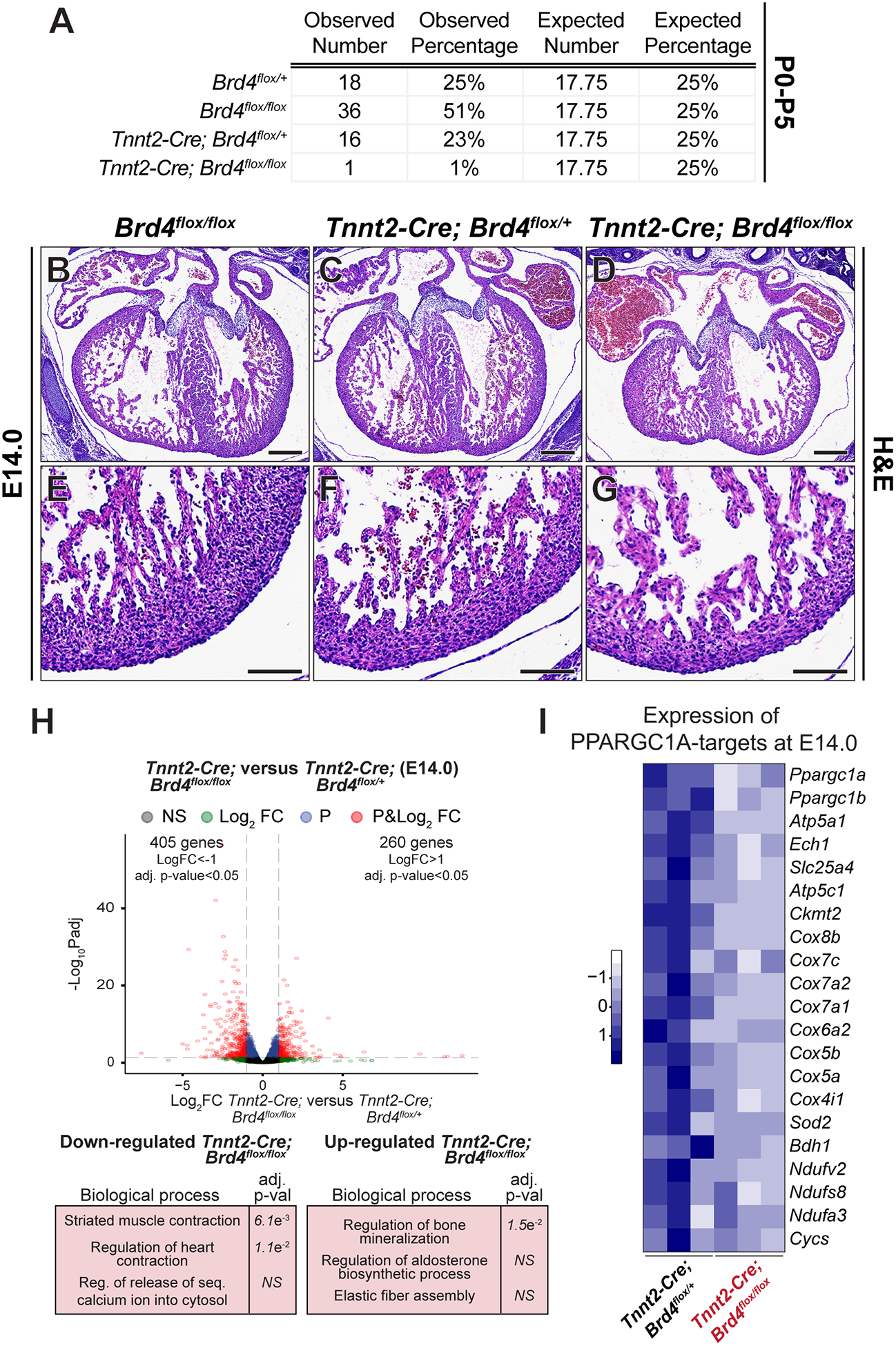
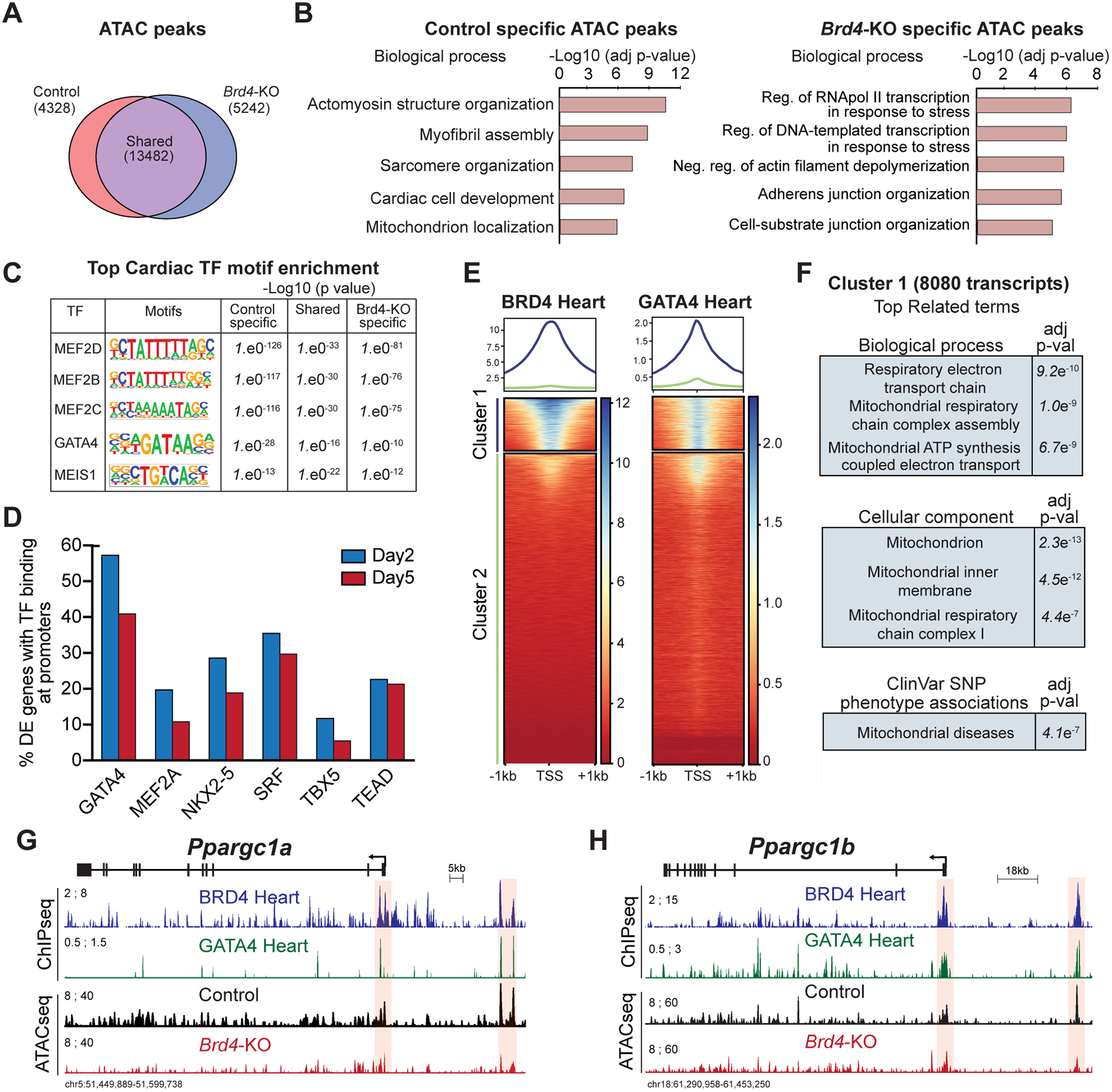
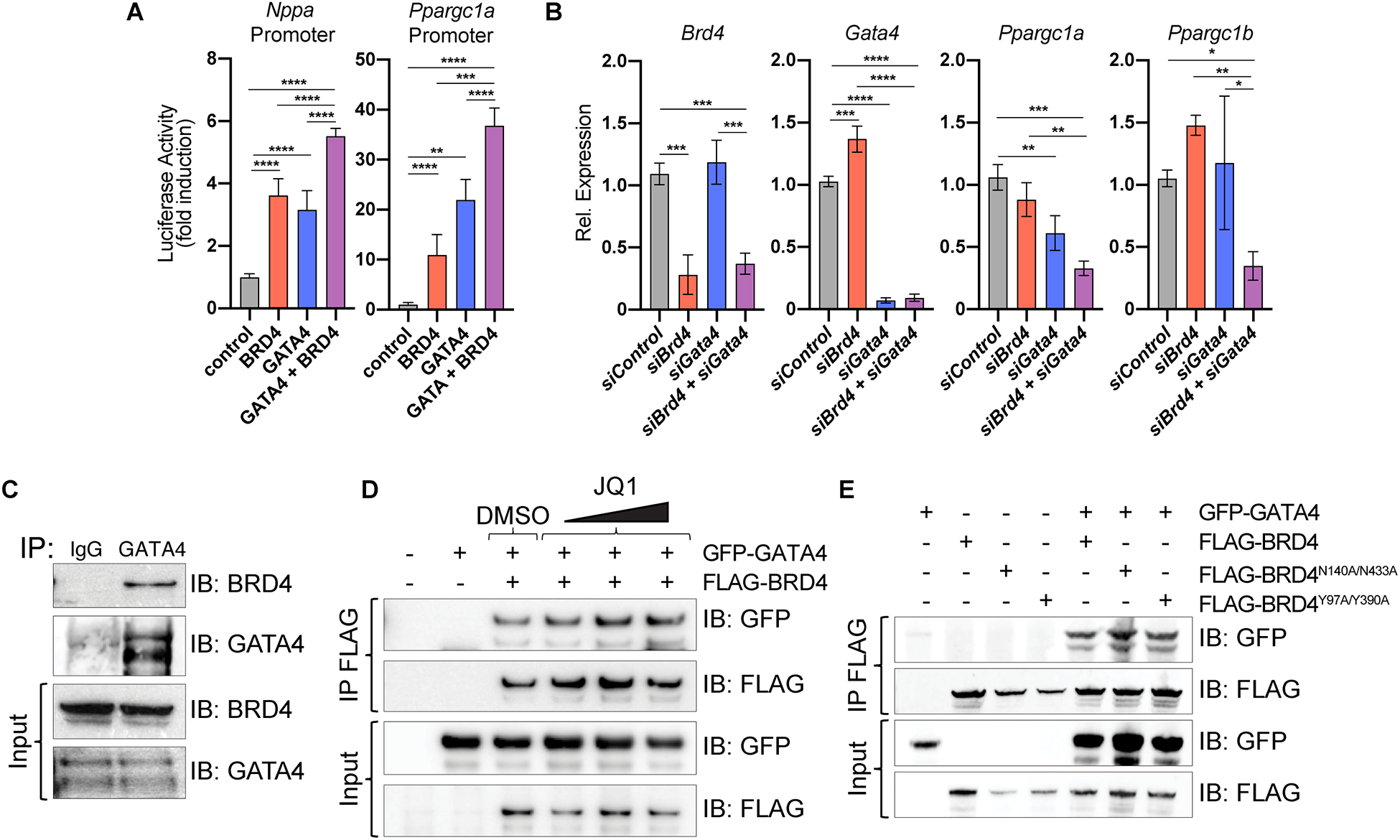
Results: Here, we show that cardiomyocyte-specific deletion of Brd4 in adult mice leads to acute deterioration of cardiac contractile function with mutant animals demonstrating a transcriptomic signature characterized by decreased expression of genes critical for mitochondrial energy production. Genome-wide occupancy data show that BRD4 enriches at many downregulated genes (including the master coactivators Ppargc1a, Ppargc1b, and their downstream targets) and preferentially colocalizes with GATA4 (GATA binding protein 4), a lineage-determining cardiac transcription factor not previously implicated in regulation of adult cardiac metabolism. BRD4 and GATA4 form an endogenous complex in cardiomyocytes and interact in a bromodomain-independent manner, revealing a new functional interaction partner for BRD4 that can direct its locus and tissue specificity.
Conclusions: These results highlight a novel role for a BRD4-GATA4 module in cooperative regulation of a cardiomyocyte-specific gene program governing bioenergetic homeostasis in the adult heart.
Keywords: epigenomics; mitochondria; myocytes, cardiac.
Figures
References
-
- Virani SS, Alonso A, Benjamin EJ, Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Chang AR, Cheng S, Delling FN, Djousse L, Elkind MSV, Ferguson JF, Fornage M, Khan SS, Kissela BM, Knutson KL, Kwan TW, Lackland DT, Lewis TT, Lichtman JH, Longenecker CT, Loop MS, Lutsey PL, Martin SS, Matsushita K, Moran AE, Mussolino ME, Perak AM, Rosamond WD, Roth GA, Sampson UKA, Satou GM, Schroeder EB, Shah SH, Shay CM, Spartano NL, Stokes A, Tirschwell DL, VanWagner LB, Tsao CW, American Heart Association Council on Epidemiology and Prevention Statistics Committee and Stroke Statistics Subcommittee. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation. 2020;141:e139–e596. - PubMed
-
- Timmis A, Townsend N, Gale CP, Torbica A, Lettino M, Petersen SE, Mossialos EA, Maggioni AP, Kazakiewicz D, May HT, De Smedt D, Flather M, Zuhlke L, Beltrame JF, Huculeci R, Tavazzi L, Hindricks G, Bax J, Casadei B, Achenbach S, Wright L, Vardas P, European Society of Cardiology. European Society of Cardiology: Cardiovascular Disease Statistics 2019. Eur Heart J. 2020;41:12–85. - PubMed
-
- Hill JA, Olson EN. Cardiac plasticity. N Engl J Med. 2008;358:1370–1380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
