

Detecting Tumor Antigen-Specific T Cells via Interaction-Dependent Fucosyl-Biotinylation
- PMID: 33096019
- PMCID: PMC7669731
- DOI: 10.1016/j.cell.2020.09.048
Detecting Tumor Antigen-Specific T Cells via Interaction-Dependent Fucosyl-Biotinylation
Abstract
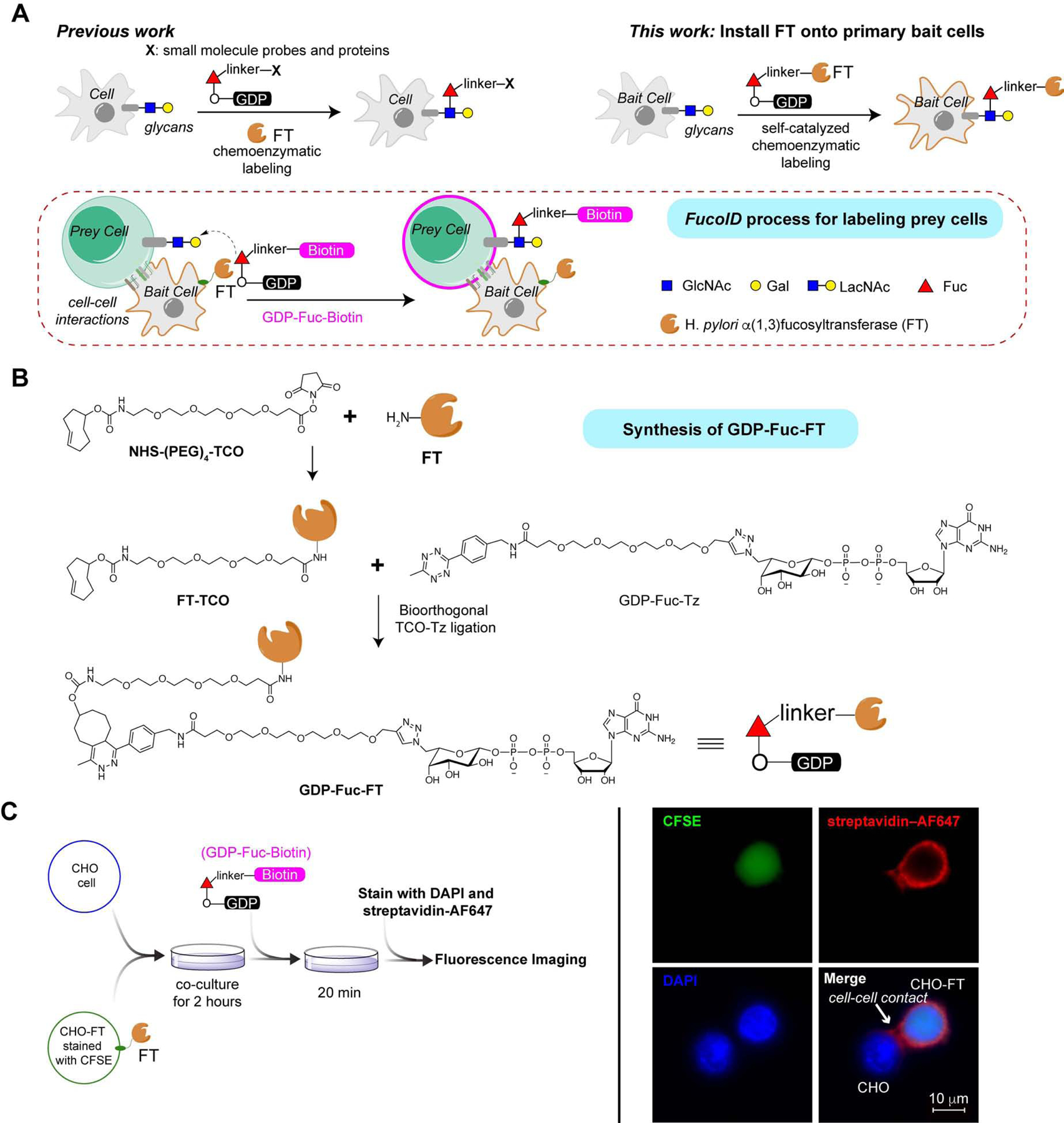
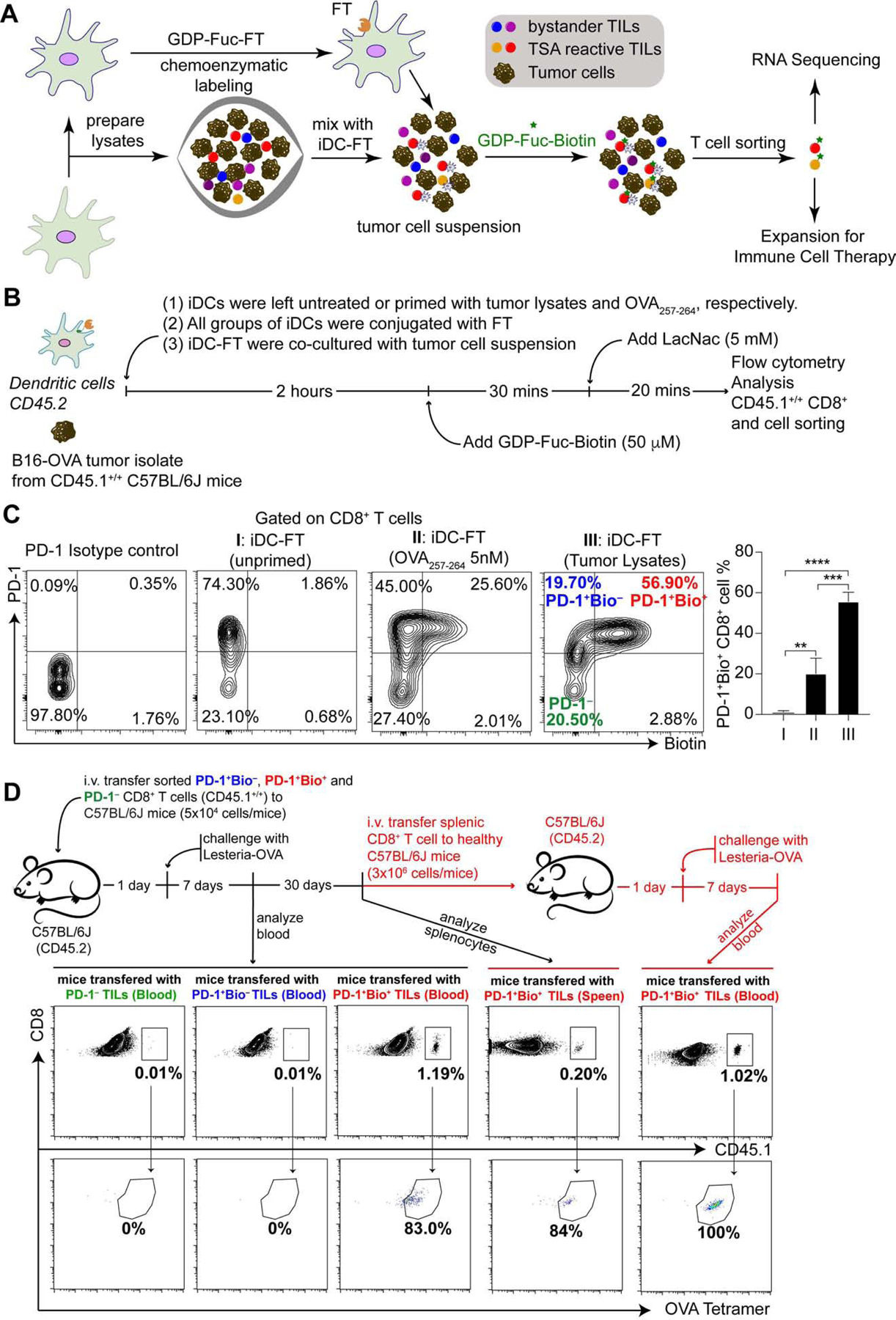
Re-activation and clonal expansion of tumor-specific antigen (TSA)-reactive T cells are critical to the success of checkpoint blockade and adoptive transfer of tumor-infiltrating lymphocyte (TIL)-based therapies. There are no reliable markers to specifically identify the repertoire of TSA-reactive T cells due to their heterogeneous composition. We introduce FucoID as a general platform to detect endogenous antigen-specific T cells for studying their biology. Through this interaction-dependent labeling approach, intratumoral TSA-reactive CD4+, CD8+ T cells, and TSA-suppressive CD4+ T cells can be detected and separated from bystander T cells based on their cell-surface enzymatic fucosyl-biotinylation. Compared to bystander TILs, TSA-reactive TILs possess a distinct T cell receptor (TCR) repertoire and unique gene features. Although exhibiting a dysfunctional phenotype, TSA-reactive CD8+ TILs possess substantial capabilities of proliferation and tumor-specific killing. Featuring genetic manipulation-free procedures and a quick turnover cycle, FucoID should have the potential of accelerating the pace of personalized cancer treatment.
Keywords: TCR sequencing; antigen-specific T cell; bystander T cell; cancer; cell-cell interaction; glycosylation; immunotherapy.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests Z.L., J.P.L., J.R.T., and P.W. are listed as inventors on a patent application filed on June 3, 2019 (U.S. Patent Application 62/856,551). All experiments were performed at Scripps Research, La Jolla, CA.
Figures





Comment in
-
Approach Enables Isolation of Tumor-Specific Antigen-Reactive T Cells.Cancer Discov. 2021 Jan;11(1):9. doi: 10.1158/2159-8290.CD-RW2020-160. Epub 2020 Nov 6. Cancer Discov. 2021. PMID: 33158846
-
Some Like It Sweet: Dendritic Cells Add Sugar to Their T(ea).Cell. 2020 Nov 12;183(4):847-849. doi: 10.1016/j.cell.2020.10.033. Cell. 2020. PMID: 33186527
References
-
- Arnaud M, Duchamp M, Bobisse S, Renaud P, Coukos G, and Harari A (2020). Biotechnologies to tackle the challenge of neoantigen identification. Curr. Opin. Biotechnol 65, 52–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
