

Collagen XII mediated cellular and extracellular mechanisms regulate establishment of tendon structure and function
- PMID: 33096204
- PMCID: PMC7870578
- DOI: 10.1016/j.matbio.2020.10.004
Collagen XII mediated cellular and extracellular mechanisms regulate establishment of tendon structure and function
Abstract
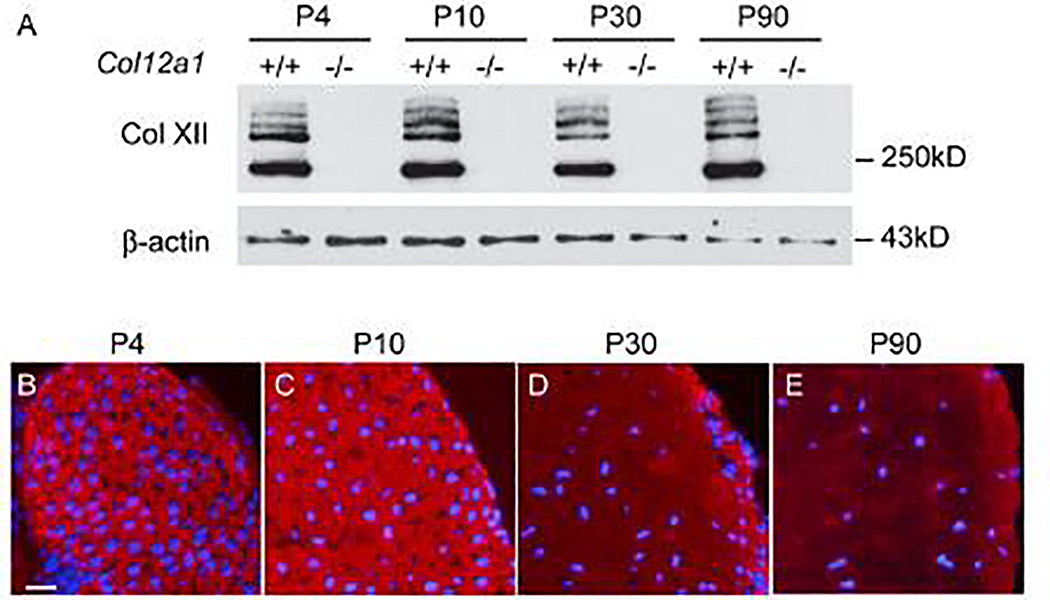
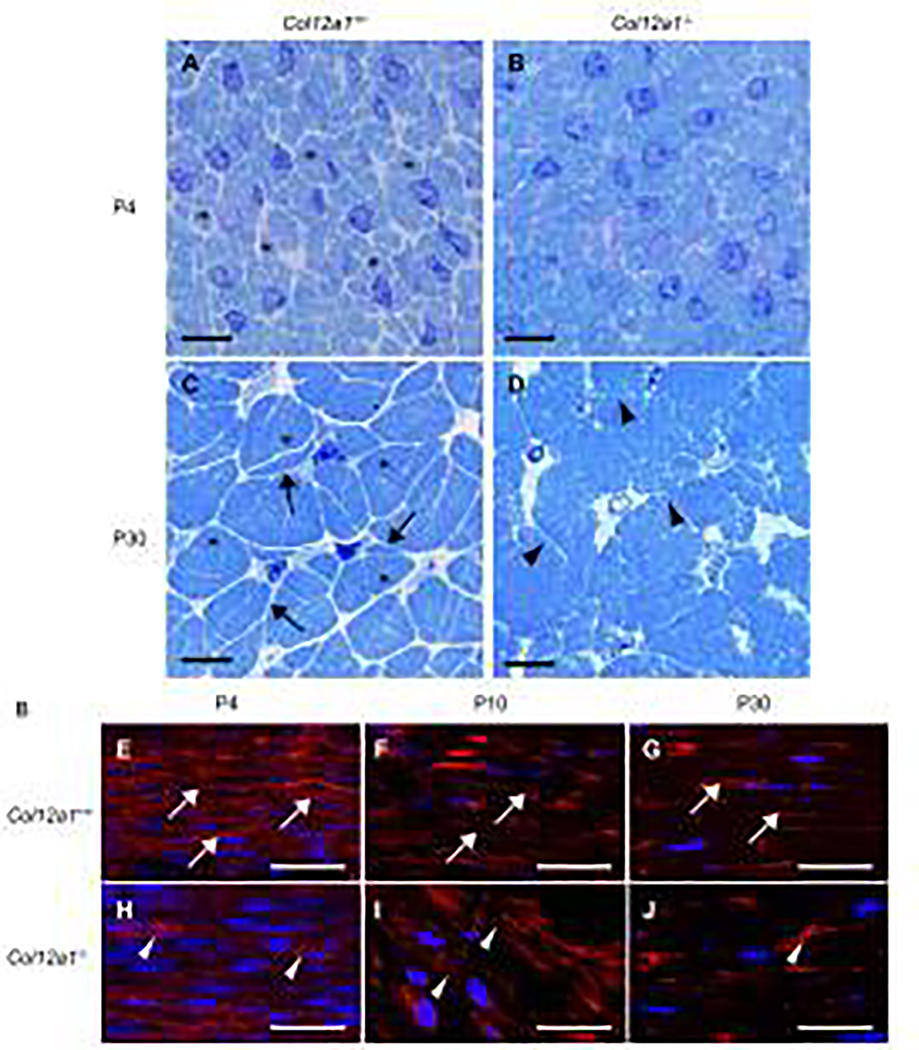
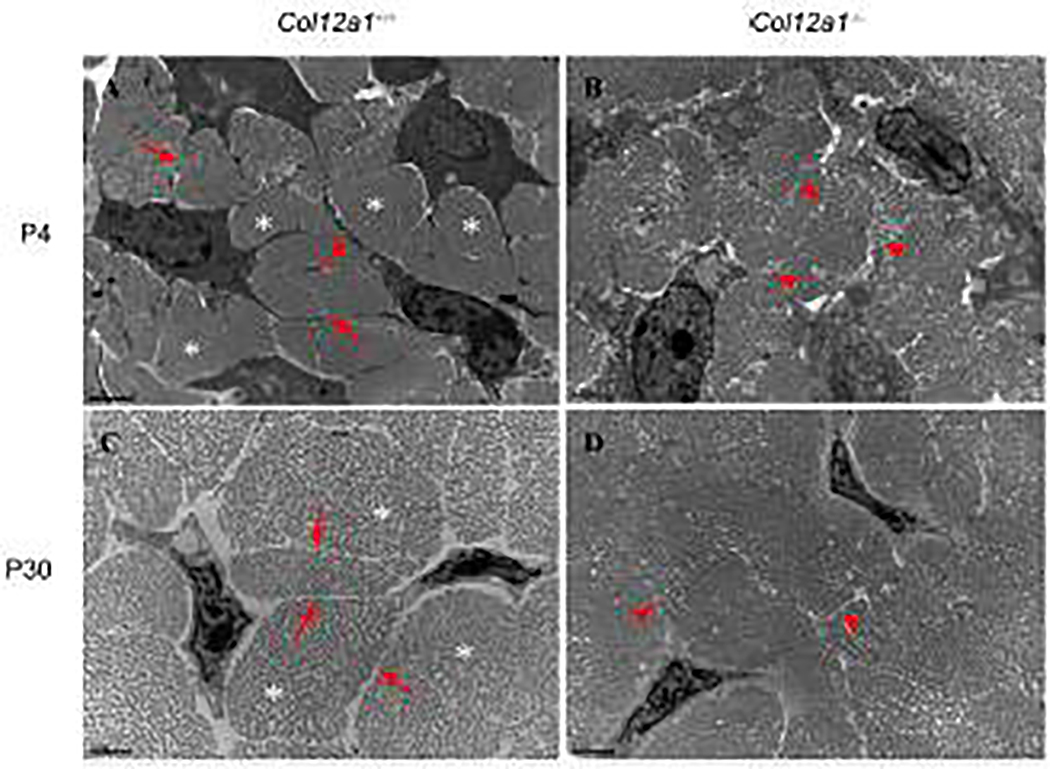
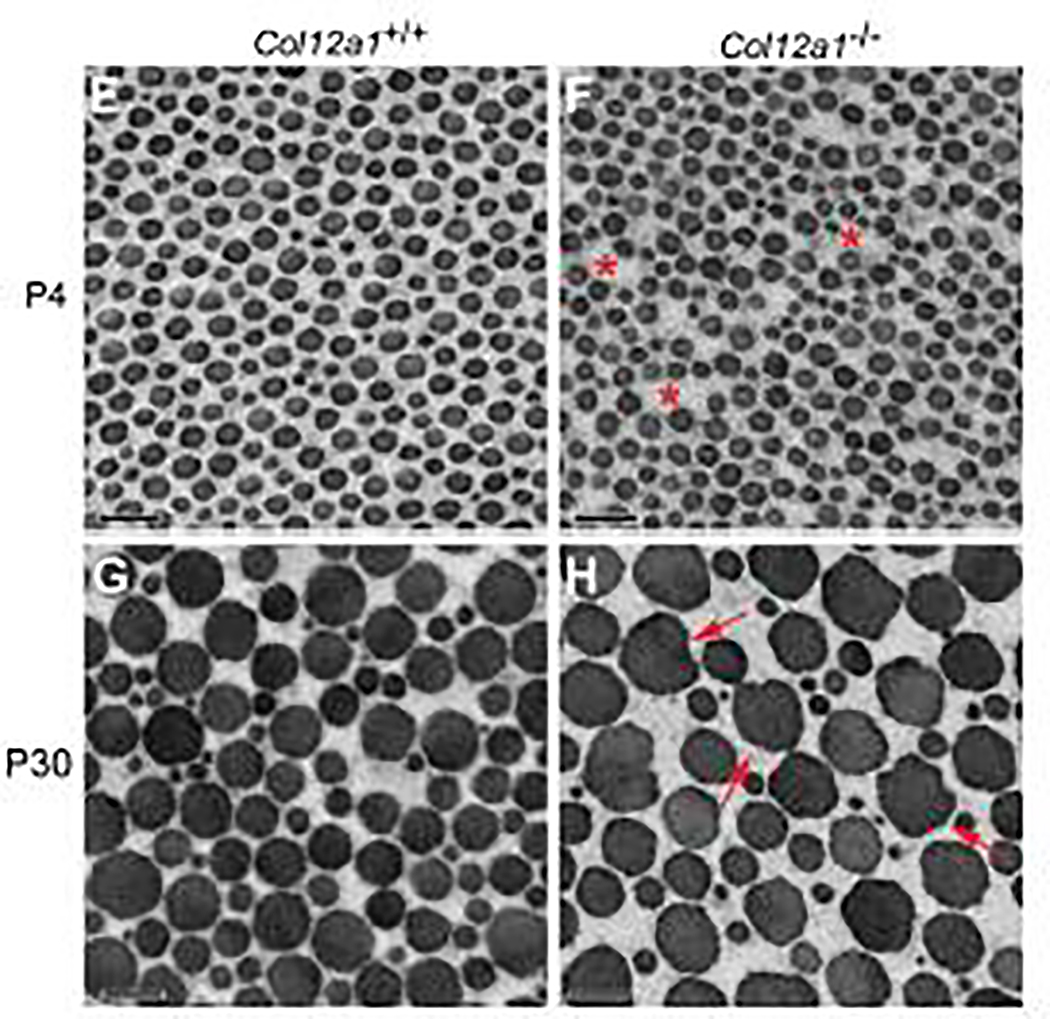
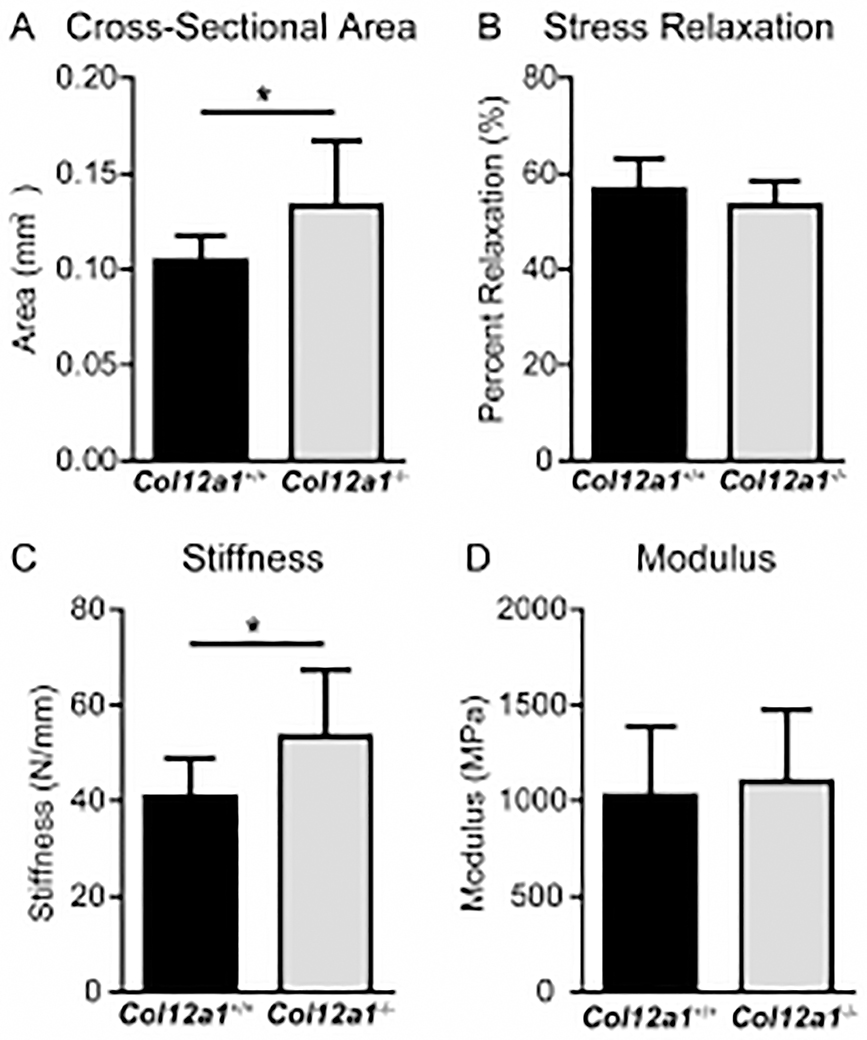
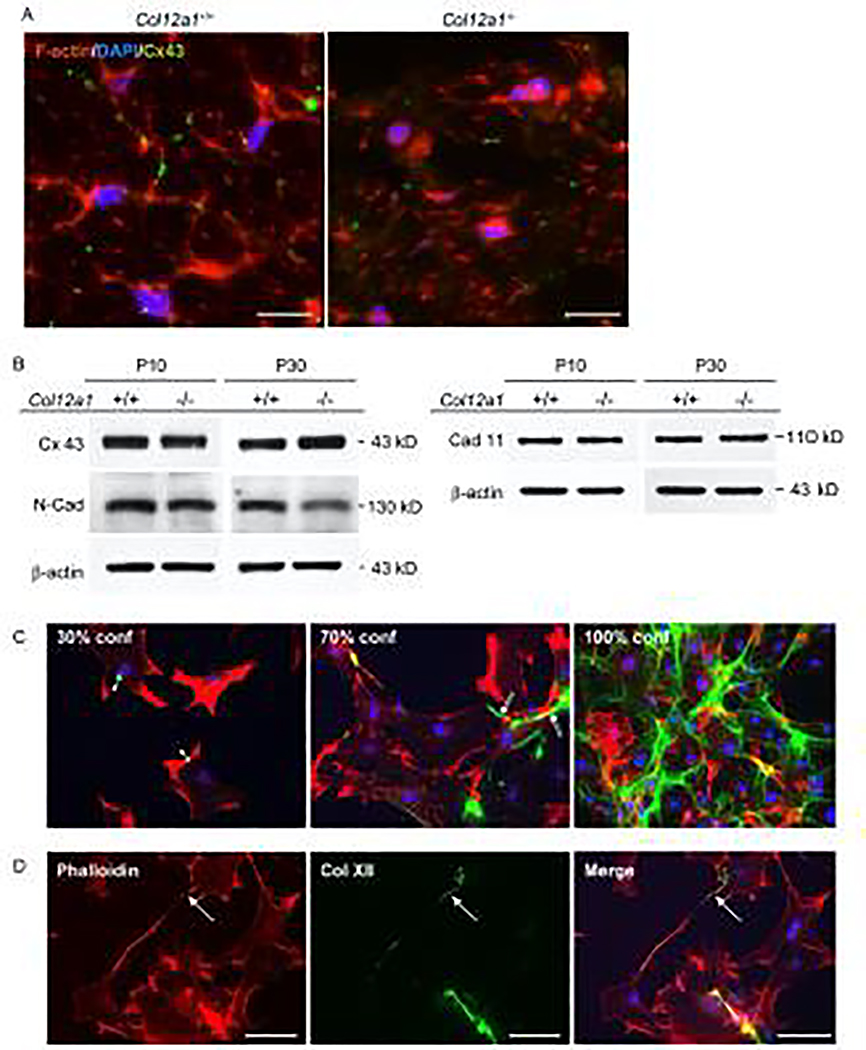
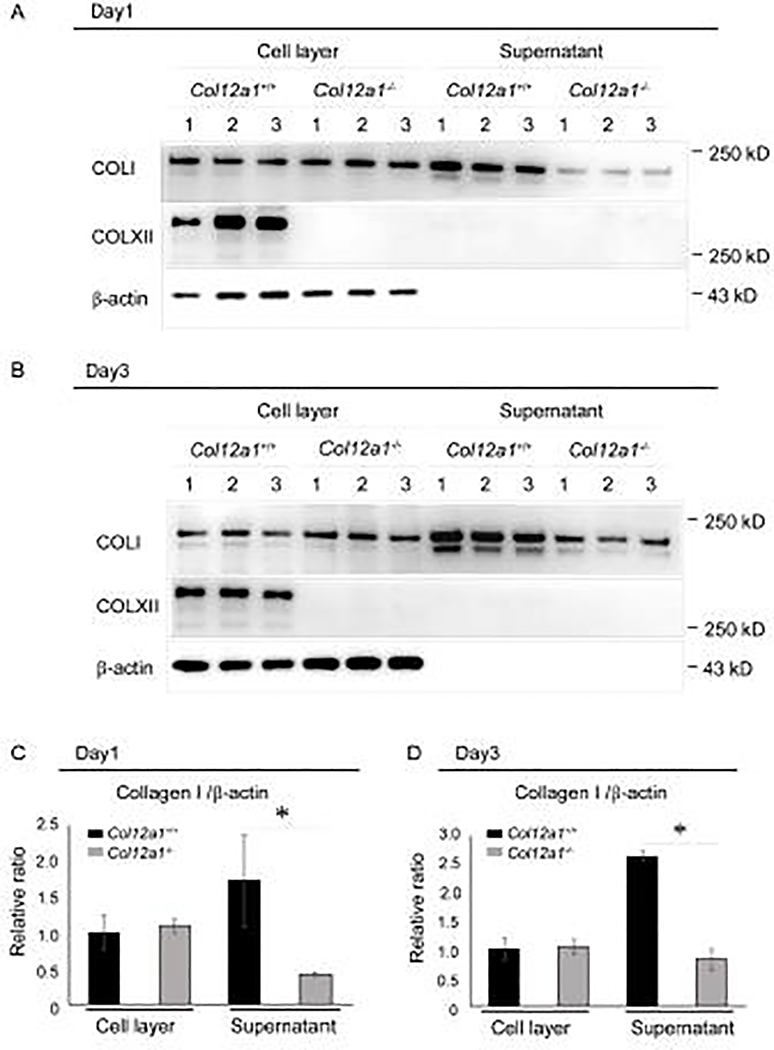
Tendons have a uniaxially aligned structure with a hierarchical organization of collagen fibrils crucial for tendon function. Collagen XII is expressed in tendons and has been implicated in the regulation of fibrillogenesis. It is a non-fibrillar collagen belonging to the Fibril-Associated Collagens with Interrupted Triple Helices (FACIT) family. Mutations in COL12A1 cause myopathic Ehlers Danlos Syndrome with a clinical phenotype involving both joints and tendons supporting critical role(s) for collagen XII in tendon development and function. Here we demonstrate the molecular function of collagen XII during tendon development using a Col12a1 null mouse model. Col12a1 deficiency altered tenocyte shape, formation of interacting cell processes, and organization resulting in impaired cell-cell communication and disruption of hierarchal structure as well as decreased tissue stiffness. Immuno-localization revealed that collagen XII accumulated on the tenocyte surface and connected adjacent tenocytes by building matrix bridges between the cells, suggesting that collagen XII regulates intercellular communication. In addition, there was a decrease in fibrillar collagen I in collagen XII deficient tenocyte cultures compared with controls suggesting collagen XII signaling specifically alters tenocyte biosynthesis. This suggests that collagen XII provides feedback to tenocytes regulating extracellular collagen I. Together, the data indicate dual roles for collagen XII in determination of tendon structure and function. Through association with fibrils it functions in fibril packing, fiber assembly and stability. In addition, collagen XII influences tenocyte organization required for assembly of higher order structure; intercellular communication necessary to coordinate long range order and feedback on tenocytes influencing collagen synthesis. Integration of both regulatory roles is required for the acquisition of hierarchal structure and mechanical properties.
Keywords: Cell–cell communication; Collagen XII; Collagen fibril assembly; Mechanical properties; Tendon; Tendon extracellular matrix assembly.
Copyright © 2020 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The author(s) declare no competing interests.
Figures
References
-
- Zhang G, Young BB, Ezura Y, Favata M, Soslowsky LJ, Chakravarti S, Birk DE, Development of tendon structure and function: Regulation of collagen fibrillogenesis, J. Musculoskelet. Neuronal Interact. 5 (2005) 5–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
