

Changes in abscisic acid metabolism in relation to the maturation of grapevine (Vitis vinifera L., cv. Mencía) somatic embryos
- PMID: 33097003
- PMCID: PMC7585196
- DOI: 10.1186/s12870-020-02701-z
Changes in abscisic acid metabolism in relation to the maturation of grapevine (Vitis vinifera L., cv. Mencía) somatic embryos
Abstract
Background: Somatic embryogenesis in grapevines is a complex process that depends on many physiological and genetic factors. One of its main limitations is the process of precocious germination of the somatic embryos in differentiation medium. This process lowers plant conversion rates from the somatic embryos, and it is probably caused by a low endogenous abscisic acid (ABA) content.
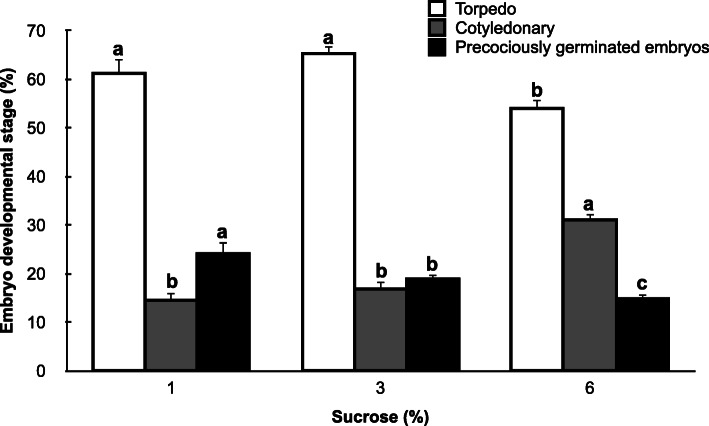
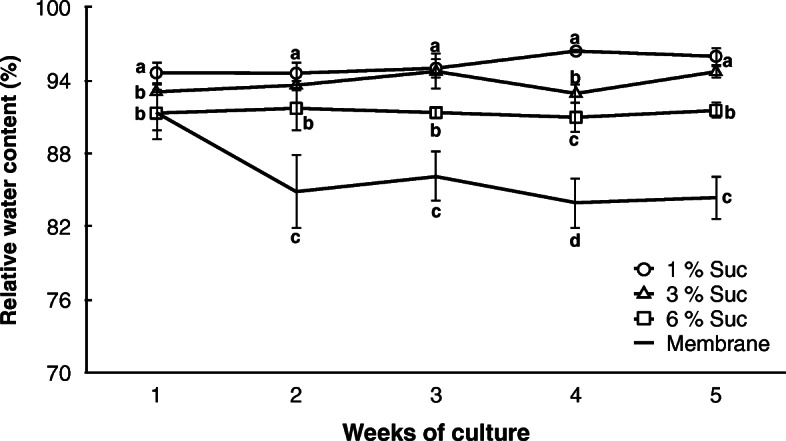
Results: Precocious germination of the somatic embryos was successfully avoided by culturing grapevine cv. Mencía embryogenic aggregates over a semipermeable membrane extended on top of the differentiation medium. The weekly analysis of the endogenous ABA and ABA-glucosyl ester (ABA-GE) contents in the aggregates showed their rapid accumulation. The expression profiles of 9-cis-epoxycarotenoid dioxygenase (VvNCED1), 8'-hydroxylase (VvHyd2), UDP-glucosyltransferase (VvUGT) and β-glucosidase (VvBG2) genes in grapevine revealed that the occurrence of a first accumulation peak of endogenous ABA in the second week of culture over the semipermeable membrane was mainly dependent on the expression of the VvNCED1 gene. A second increase in the endogenous ABA content was observed in the fourth week of culture. At this point in the culture, our results suggest that of those genes involved in ABA accumulation, one (VvNCED1) was repressed, while another (VvBG2) was activated. Similarly, of those genes related to a reduction in ABA levels, one (VvUGT) was repressed while another (VvHyd2) was activated. The relative expression level of the VvNCED1 gene in embryogenic aggregates cultured under the same conditions and treated with exogenous ABA revealed the significant downregulation of this gene.
Conclusions: Our results demonstrated the involvement of ABA metabolism in the control of the maturation of grapevine somatic embryos cultured over a semipermeable membrane and two important control points for their endogenous ABA levels. Thus, subtle differences in the expression of the antagonistic genes that control ABA synthesis and degradation could be responsible for the final level of ABA during the maturation of grapevine somatic embryos in vitro. In addition, the treatment of somatic embryos with exogenous ABA suggested the feedback-based control of the expression of the VvNCED1 gene by ABA during the maturation of grapevine somatic embryos.
Keywords: ABA metabolism; Cellulose semipermeable membrane; Gene expression; Reference genes; Somatic embryogenesis; qPCR.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





References
-
- Gray DJ, Compton ME. Chapter 20 - grape somatic embryo dormancy and quiescence: potential of dehydrated synthetic seeds for germplasm conservation. In: Redenbaugh K, editor. Synseeds: applications of synthetic seeds to crop improvement. Boca Raton: CRC Press; 1993. pp. 367–379.
-
- Senaratna T, McKersie BD, Bowley SR. Desiccation tolerance of alfalfa (Medicago sativa L.) somatic embryos - influence of abscisic-acid, stress pretreatments and drying rates. Plant Sci. 1989;65:253–259. doi: 10.1016/0168-9452(89)90072-1. - DOI
-
- Senaratna T, McKersie BD, Bowley SR. Artificial seeds of alfalfa (Medicago sativa L.). induction of desiccation tolerance in somatic embryos. In Vitro Cell Dev Bio Plant. 1990;26:85–90. doi: 10.1007/BF02624160. - DOI
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
