

Differential Short-Term Plasticity of PV and SST Neurons Accounts for Adaptation and Facilitation of Cortical Neurons to Auditory Tones
- PMID: 33097639
- PMCID: PMC7687067
- DOI: 10.1523/JNEUROSCI.0686-20.2020
Differential Short-Term Plasticity of PV and SST Neurons Accounts for Adaptation and Facilitation of Cortical Neurons to Auditory Tones
Abstract
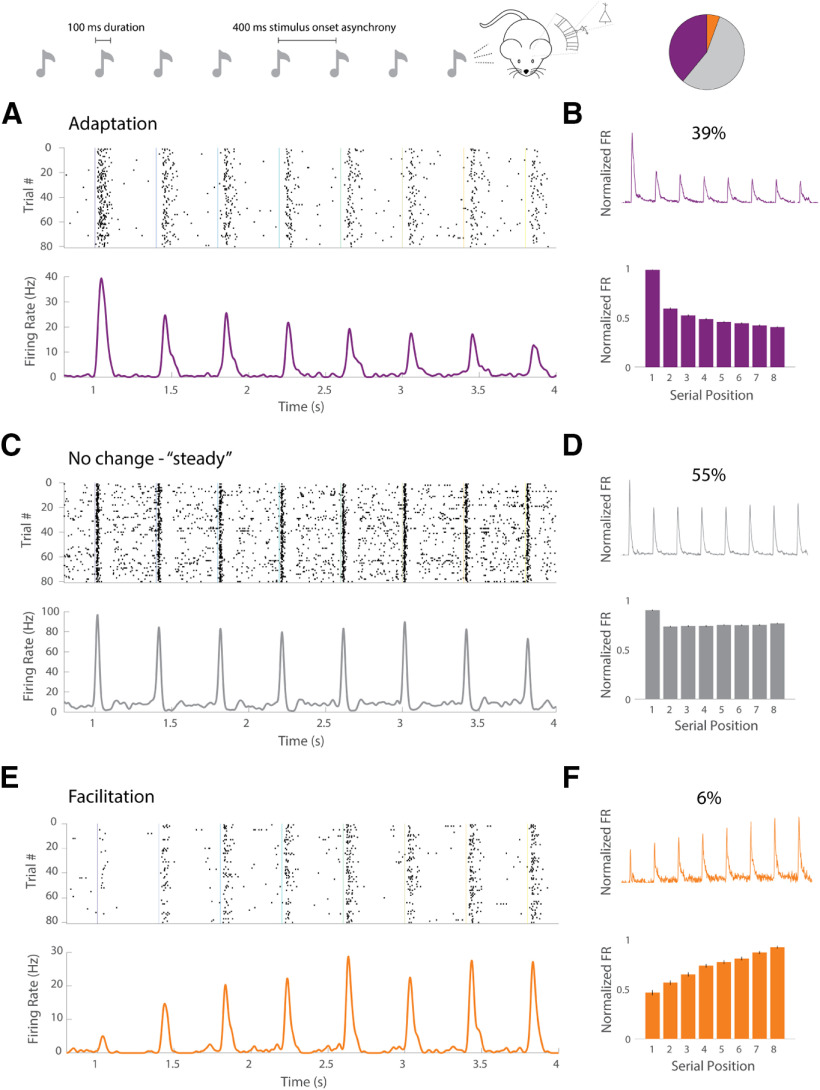
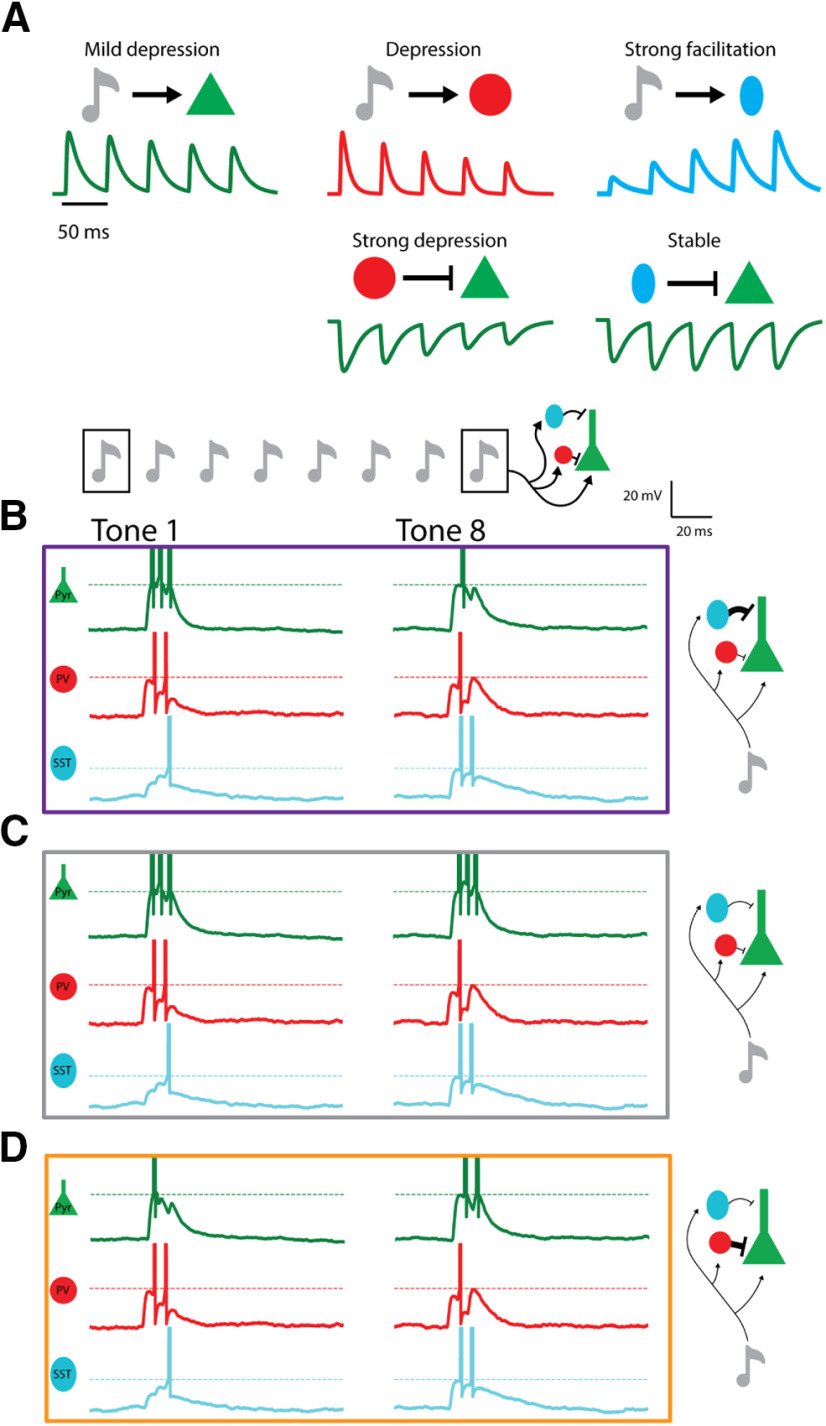
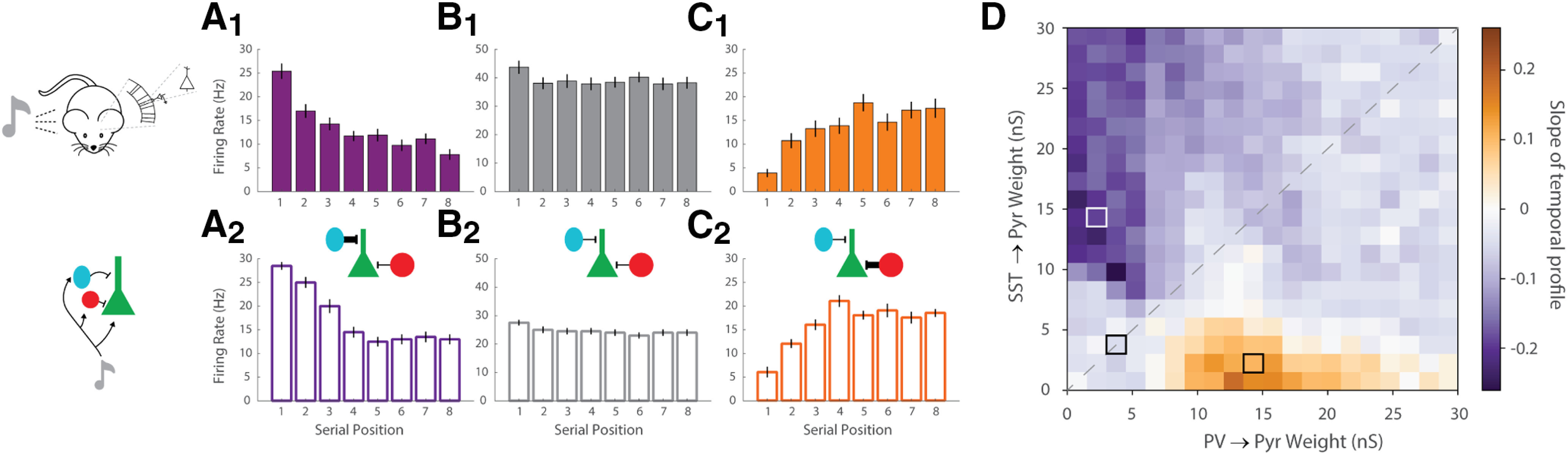
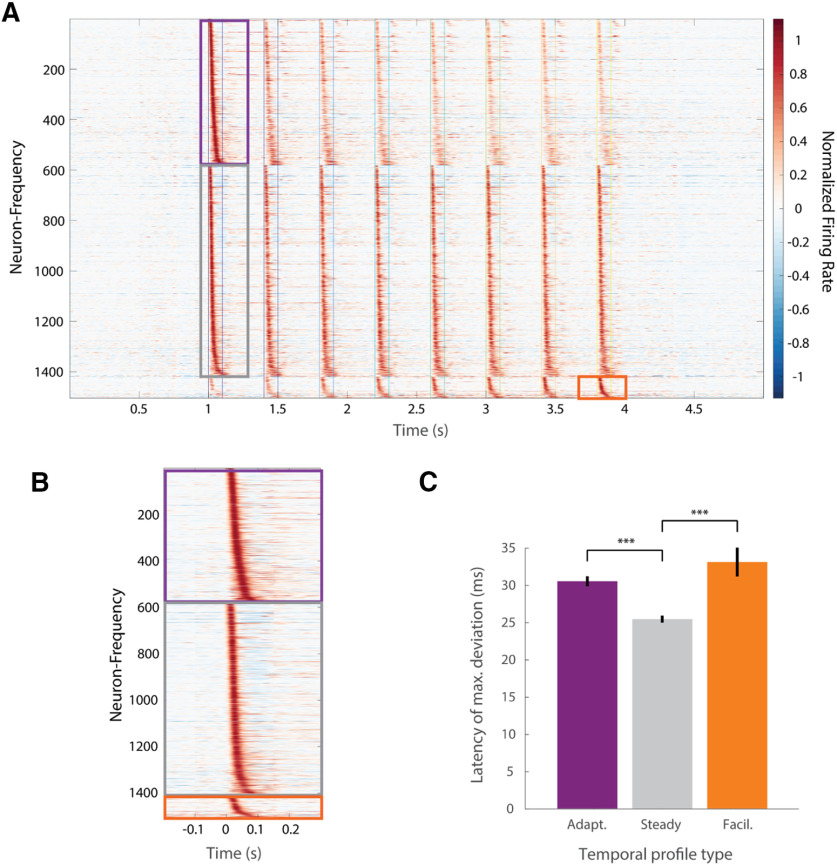
Cortical responses to sensory stimuli are strongly modulated by temporal context. One of the best studied examples of such modulation is sensory adaptation. We first show that in response to repeated tones pyramidal (Pyr) neurons in male mouse auditory cortex (A1) exhibit facilitating and stable responses, in addition to adapting responses. To examine the potential mechanisms underlying these distinct temporal profiles, we developed a reduced spiking model of sensory cortical circuits that incorporated the signature short-term synaptic plasticity (STP) profiles of the inhibitory parvalbumin (PV) and somatostatin (SST) interneurons. The model accounted for all three temporal response profiles as the result of dynamic changes in excitatory/inhibitory balance produced by STP, primarily through shifts in the relative latency of Pyr and inhibitory neurons. Transition between the three response profiles was possible by changing the strength of the inhibitory PV→Pyr and SST→Pyr synapses. The model predicted that a unit's latency would be related to its temporal profile. Consistent with this prediction, the latency of stable units was significantly shorter than that of adapting and facilitating units. Furthermore, because of the history-dependence of STP the model generated a paradoxical prediction: that inactivation of inhibitory neurons during one tone would decrease the response of A1 neurons to a subsequent tone. Indeed, we observed that optogenetic inactivation of PV neurons during one tone counterintuitively decreased the spiking of Pyr neurons to a subsequent tone 400 ms later. These results provide evidence that STP is critical to temporal context-dependent responses in the sensory cortex.SIGNIFICANCE STATEMENT Our perception of speech and music depends strongly on temporal context, i.e., the significance of a stimulus depends on the preceding stimuli. Complementary neural mechanisms are needed to sometimes ignore repetitive stimuli (e.g., the tic of a clock) or detect meaningful repetition (e.g., consecutive tones in Morse code). We modeled a neural circuit that accounts for diverse experimentally-observed response profiles in auditory cortex (A1) neurons, based on known forms of short-term synaptic plasticity (STP). Whether the simulated circuit reduced, maintained, or enhanced its response to repeated tones depended on the relative dominance of two different types of inhibitory cells. The model made novel predictions that were experimentally validated. Results define an important role for STP in temporal context-dependent perception.
Keywords: adaptation; auditory cortex; parvalbumin; short-term synaptic plasticity; somatostatin; temporal.
Copyright © 2020 the authors.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases