

Insights into Lysosomal PI(3,5)P2 Homeostasis from a Structural-Biochemical Analysis of the PIKfyve Lipid Kinase Complex
- PMID: 33098764
- PMCID: PMC7962480
- DOI: 10.1016/j.molcel.2020.10.003
Insights into Lysosomal PI(3,5)P2 Homeostasis from a Structural-Biochemical Analysis of the PIKfyve Lipid Kinase Complex
Abstract
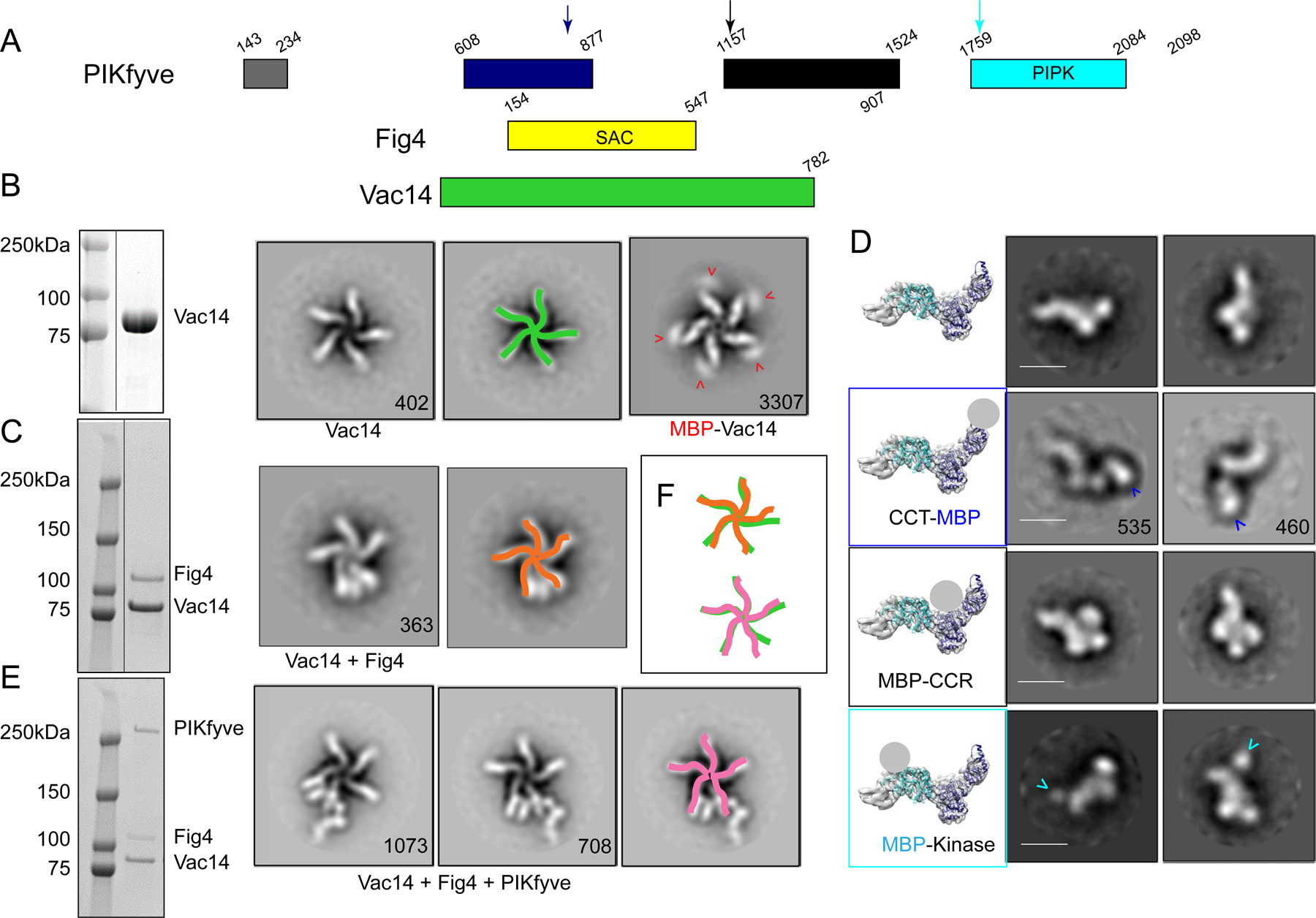
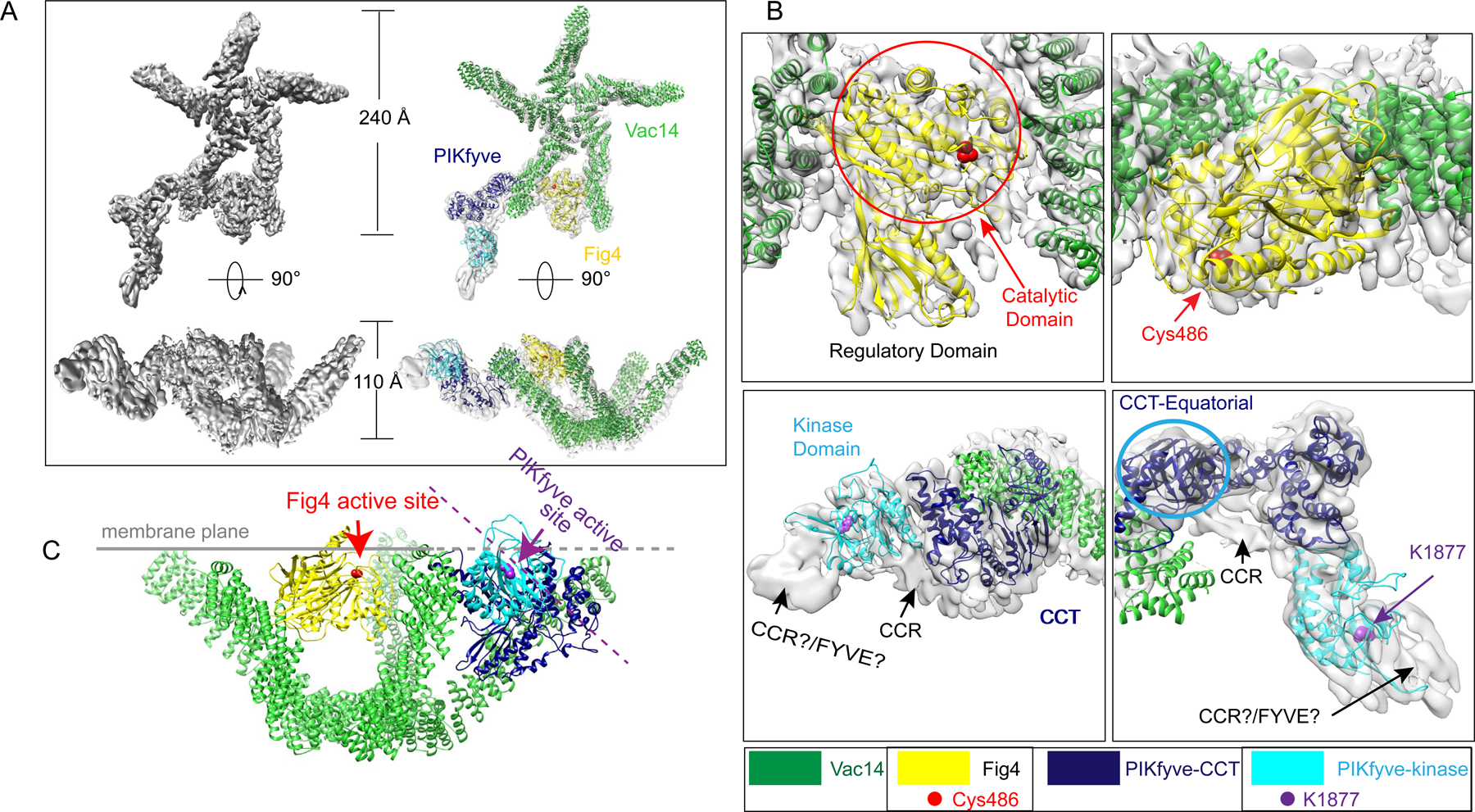
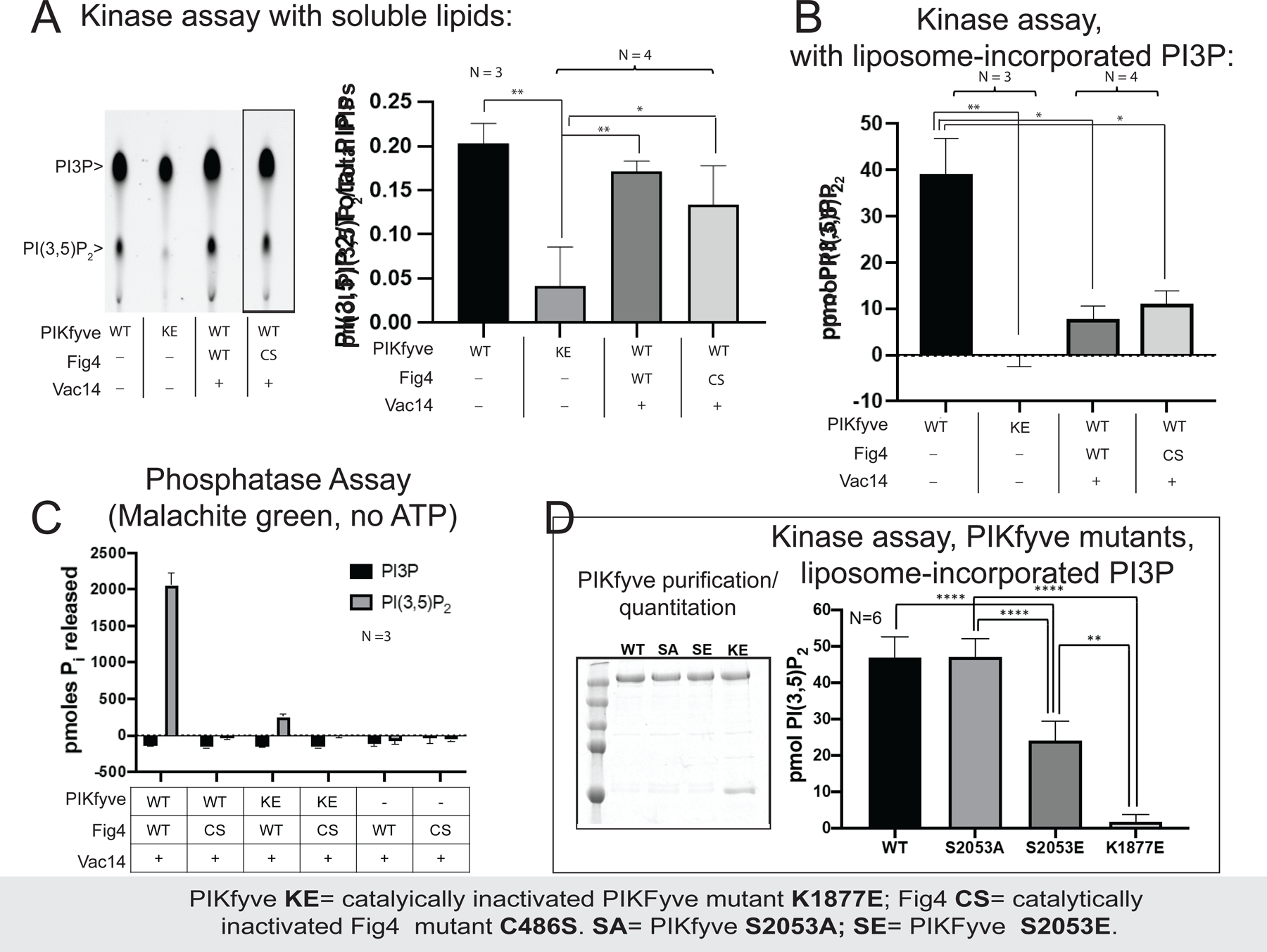
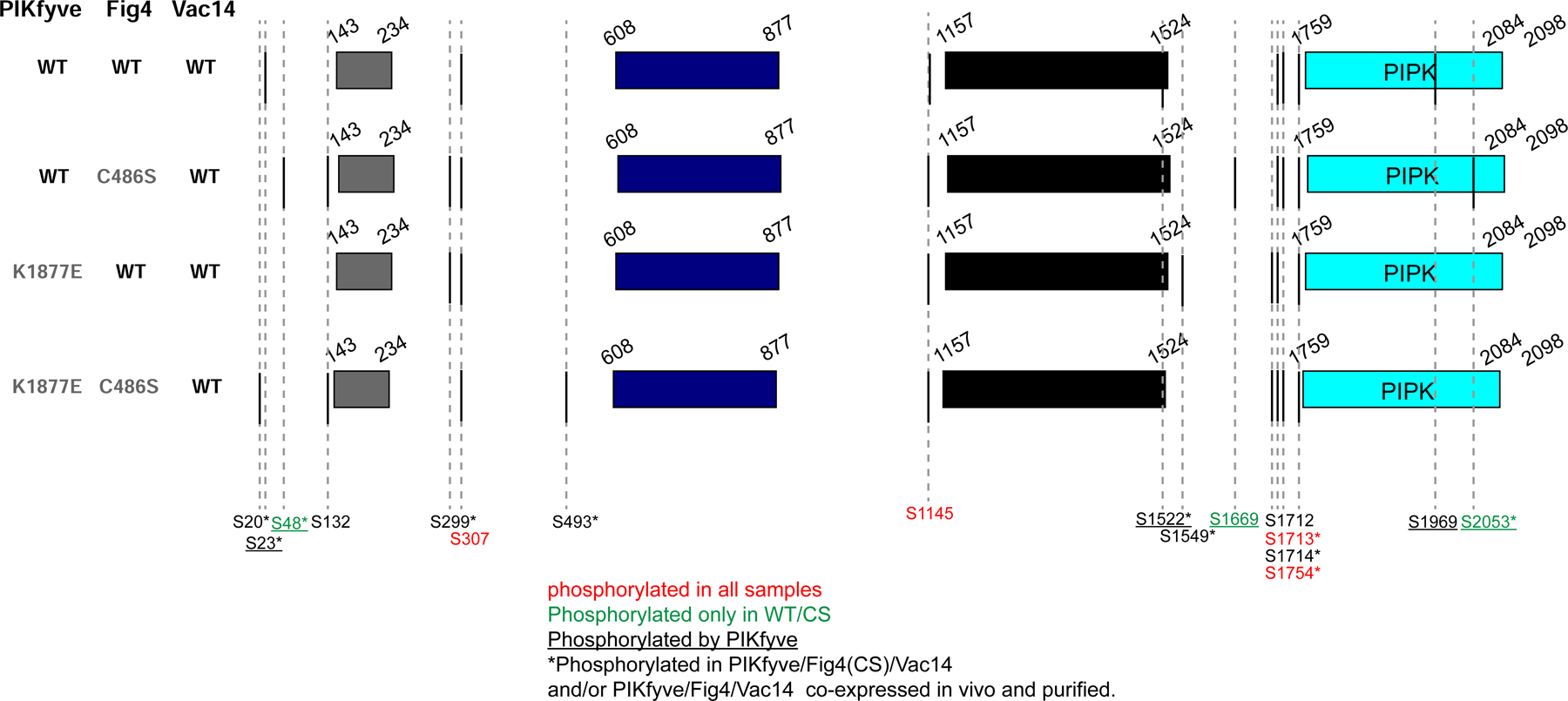
The phosphoinositide PI(3,5)P2, generated exclusively by the PIKfyve lipid kinase complex, is key for lysosomal biology. Here, we explore how PI(3,5)P2 levels within cells are regulated. We find the PIKfyve complex comprises five copies of the scaffolding protein Vac14 and one copy each of the lipid kinase PIKfyve, generating PI(3,5)P2 from PI3P and the lipid phosphatase Fig4, reversing the reaction. Fig4 is active as a lipid phosphatase in the ternary complex, whereas PIKfyve within the complex cannot access membrane-incorporated phosphoinositides due to steric constraints. We find further that the phosphoinositide-directed activities of both PIKfyve and Fig4 are regulated by protein-directed activities within the complex. PIKfyve autophosphorylation represses its lipid kinase activity and stimulates Fig4 lipid phosphatase activity. Further, Fig4 is also a protein phosphatase acting on PIKfyve to stimulate its lipid kinase activity, explaining why catalytically active Fig4 is required for maximal PI(3,5)P2 production by PIKfyve in vivo.
Keywords: lipid kinase; lipid phosphatase; phosphoinositide homeostasis.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
