

Non-Cell-Autonomous Mechanisms in Radial Projection Neuron Migration in the Developing Cerebral Cortex
- PMID: 33102480
- PMCID: PMC7545535
- DOI: 10.3389/fcell.2020.574382
Non-Cell-Autonomous Mechanisms in Radial Projection Neuron Migration in the Developing Cerebral Cortex
Abstract
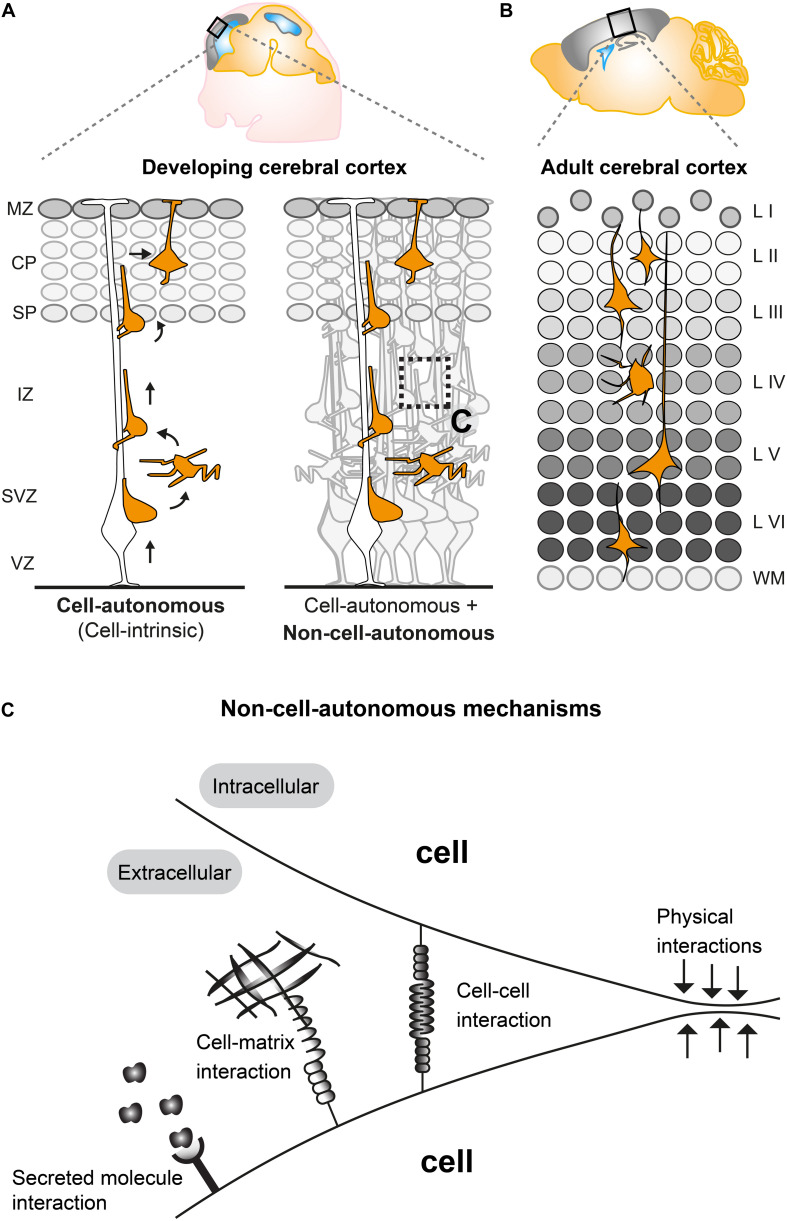
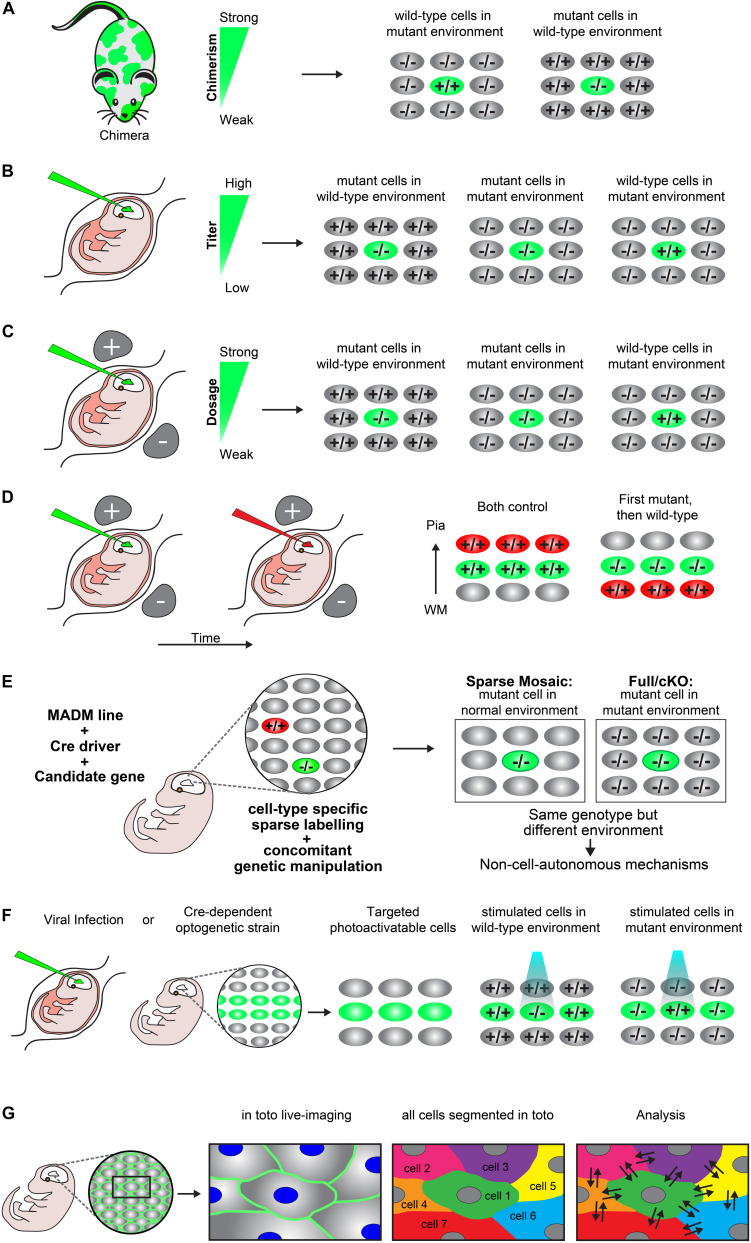
Concerted radial migration of newly born cortical projection neurons, from their birthplace to their final target lamina, is a key step in the assembly of the cerebral cortex. The cellular and molecular mechanisms regulating the specific sequential steps of radial neuronal migration in vivo are however still unclear, let alone the effects and interactions with the extracellular environment. In any in vivo context, cells will always be exposed to a complex extracellular environment consisting of (1) secreted factors acting as potential signaling cues, (2) the extracellular matrix, and (3) other cells providing cell-cell interaction through receptors and/or direct physical stimuli. Most studies so far have described and focused mainly on intrinsic cell-autonomous gene functions in neuronal migration but there is accumulating evidence that non-cell-autonomous-, local-, systemic-, and/or whole tissue-wide effects substantially contribute to the regulation of radial neuronal migration. These non-cell-autonomous effects may differentially affect cortical neuron migration in distinct cellular environments. However, the cellular and molecular natures of such non-cell-autonomous mechanisms are mostly unknown. Furthermore, physical forces due to collective migration and/or community effects (i.e., interactions with surrounding cells) may play important roles in neocortical projection neuron migration. In this concise review, we first outline distinct models of non-cell-autonomous interactions of cortical projection neurons along their radial migration trajectory during development. We then summarize experimental assays and platforms that can be utilized to visualize and potentially probe non-cell-autonomous mechanisms. Lastly, we define key questions to address in the future.
Keywords: cerebral cortex; neurodevelopmental migration disorders; non-cell-autonomous mechanisms; radial projection neuron migration; single cell analysis.
Copyright © 2020 Hansen and Hippenmeyer.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources