

Metabolome Changes during In Vivo Red Cell Aging Reveal Disruption of Key Metabolic Pathways
- PMID: 33103072
- PMCID: PMC7575880
- DOI: 10.1016/j.isci.2020.101630
Metabolome Changes during In Vivo Red Cell Aging Reveal Disruption of Key Metabolic Pathways
Abstract
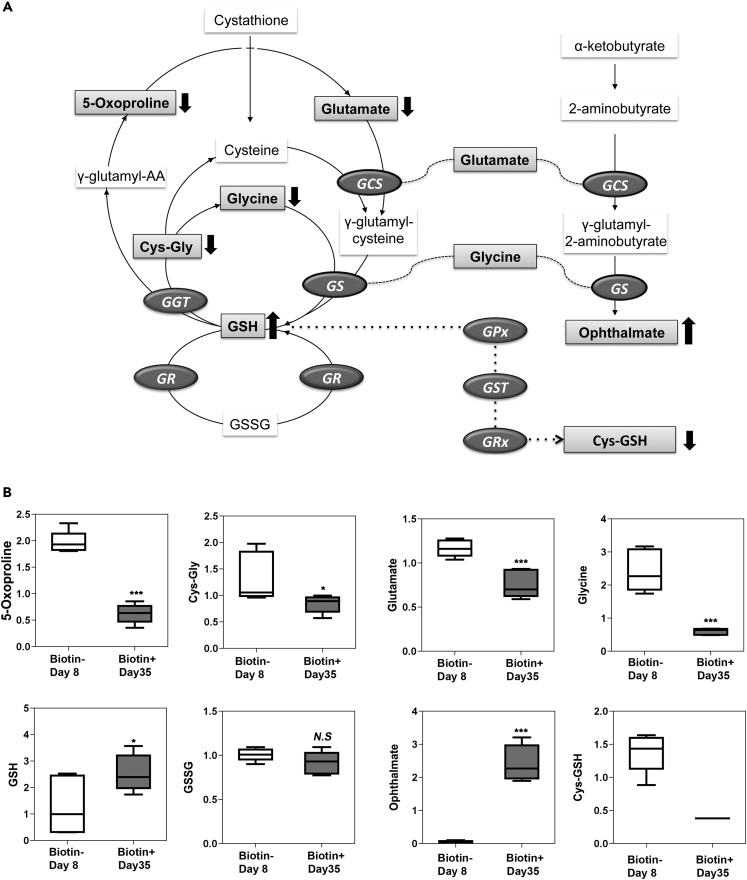
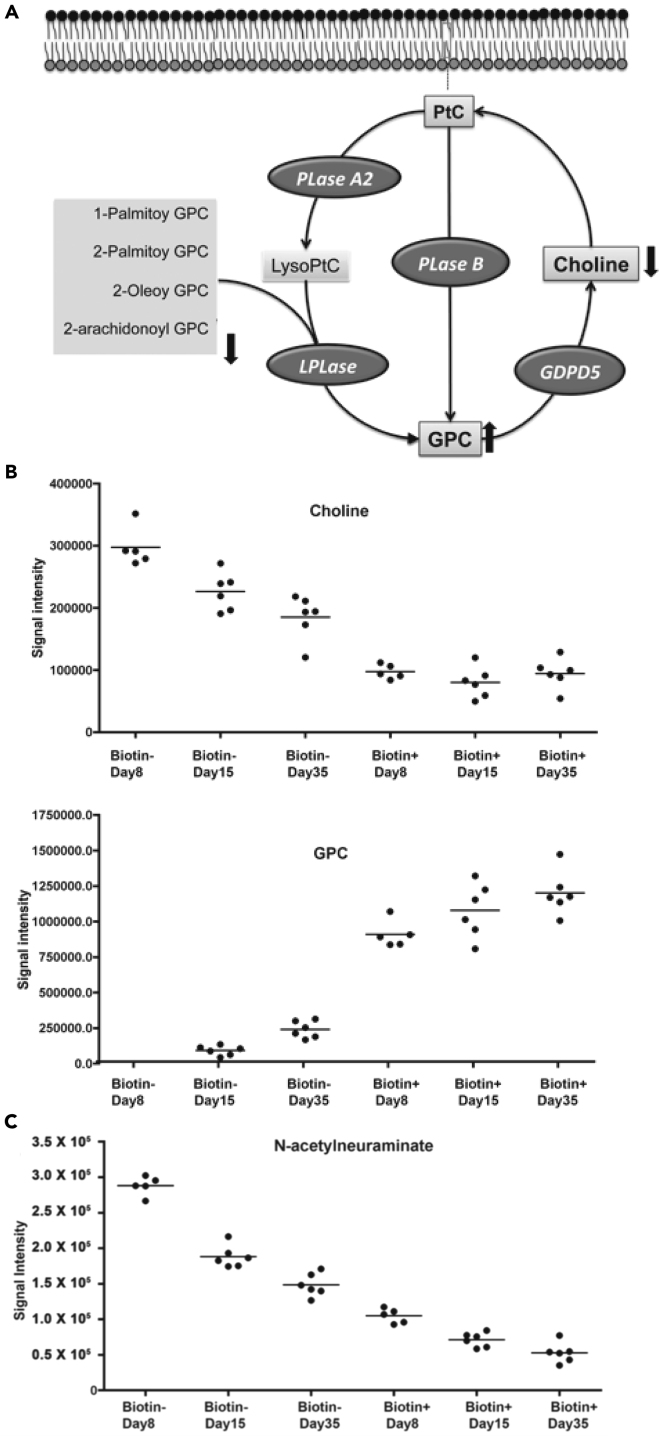
Understanding the mechanisms for cellular aging is a fundamental question in biology. Normal red blood cells (RBCs) survive for approximately 100 days, and their survival is likely limited by functional decline secondary to cumulative damage to cell constituents, which may be reflected in altered metabolic capabilities. To investigate metabolic changes during in vivo RBC aging, labeled cell populations were purified at intervals and assessed for abundance of metabolic intermediates using mass spectrometry. A total of 167 metabolites were profiled and quantified from cell populations of defined ages. Older RBCs maintained ATP and redox charge states at the cost of altered activity of enzymatic pathways. Time-dependent changes were identified in metabolites related to maintenance of the redox state and membrane structure. These findings illuminate the differential metabolic pathway usage associated with normal cellular aging and identify potential biomarkers to determine average RBC age and rates of RBC turnover from a single blood sample.
Keywords: Medical Biochemistry; Metabolomics.
© 2020 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







Similar articles
-
Identified metabolic signature for assessing red blood cell unit quality is associated with endothelial damage markers and clinical outcomes.Transfusion. 2016 Apr;56(4):852-62. doi: 10.1111/trf.13460. Epub 2016 Jan 8. Transfusion. 2016. PMID: 26749434
-
Time-Course Investigation of Small Molecule Metabolites in MAP-Stored Red Blood Cells Using UPLC-QTOF-MS.Molecules. 2018 Apr 16;23(4):923. doi: 10.3390/molecules23040923. Molecules. 2018. PMID: 29659551 Free PMC article.
-
Parabiosis Incompletely Reverses Aging-Induced Metabolic Changes and Oxidant Stress in Mouse Red Blood Cells.Nutrients. 2019 Jun 14;11(6):1337. doi: 10.3390/nu11061337. Nutrients. 2019. PMID: 31207887 Free PMC article.
-
Exercise, training and red blood cell turnover.Sports Med. 1995 Jan;19(1):9-31. doi: 10.2165/00007256-199519010-00002. Sports Med. 1995. PMID: 7740249 Review.
-
Challenges for red blood cell biomarker discovery through proteomics.Biochim Biophys Acta. 2014 May;1844(5):1003-10. doi: 10.1016/j.bbapap.2013.10.002. Epub 2013 Oct 12. Biochim Biophys Acta. 2014. PMID: 24129076 Review.
Cited by
-
Red Blood Cell Omics and Machine Learning in Transfusion Medicine: Singularity Is Near.Transfus Med Hemother. 2023 Mar 8;50(3):174-183. doi: 10.1159/000529744. eCollection 2023 Jun. Transfus Med Hemother. 2023. PMID: 37434999 Free PMC article. Review.
-
Inductively-Coupled Plasma Mass Spectrometry-Novel Insights From an Old Technology Into Stressed Red Blood Cell Physiology.Front Physiol. 2022 Feb 7;13:828087. doi: 10.3389/fphys.2022.828087. eCollection 2022. Front Physiol. 2022. PMID: 35197866 Free PMC article.
-
Drug transporters OAT1 and OAT3 have specific effects on multiple organs and gut microbiome as revealed by contextualized metabolic network reconstructions.Sci Rep. 2022 Oct 31;12(1):18308. doi: 10.1038/s41598-022-21091-w. Sci Rep. 2022. PMID: 36316339 Free PMC article.
-
The Role of Ergothioneine in Red Blood Cell Biology: A Review and Perspective.Antioxidants (Basel). 2024 Jun 13;13(6):717. doi: 10.3390/antiox13060717. Antioxidants (Basel). 2024. PMID: 38929156 Free PMC article. Review.
-
RBC-GEM: A genome-scale metabolic model for systems biology of the human red blood cell.PLoS Comput Biol. 2025 Mar 12;21(3):e1012109. doi: 10.1371/journal.pcbi.1012109. eCollection 2025 Mar. PLoS Comput Biol. 2025. PMID: 40072998 Free PMC article.
References
-
- Akanmu D., Cecchini R., Aruoma O.I., Halliwell B. The antioxidant action of ergothioneine. Arch. Biochem. Biophys. 1991;288:10–16. - PubMed
-
- Aminoff D., Bruegge W.F., Bell W.C., Sarpolis K., Williams R. Role of sialic acid in survival of erythrocytes in the circulation: interaction of neuraminidase-treated and untreated erythrocytes with spleen and liver at the cellular level. Proc. Natl. Acad. Sci. U S A. 1977;74:1521–1524. - PMC - PubMed
-
- Arnold L.W., Haughton G. Autoantibodies to phosphatidylcholine. The murine antibromelain RBC response. Ann. N Y Acad. Sci. 1992;651:354–359. - PubMed
-
- Aruoma O.I., Whiteman M., England T.G., Halliwell B. Antioxidant action of ergothioneine: assessment of its ability to scavenge peroxynitrite. Biochem. Biophys. Res. Commun. 1997;231:389–391. - PubMed
-
- Bacher P., Giersiefer S., Bach M., Fork C., Schomig E., Grundemann D. Substrate discrimination by ergothioneine transporter SLC22A4 and carnitine transporter SLC22A5: gain-of-function by interchange of selected amino acids. Biochim. Biophys. Acta. 2009;1788:2594–2602. - PubMed
