

Mitochondrial Protein Synthesis Is Essential for Terminal Differentiation of CD45- TER119-Erythroid and Lymphoid Progenitors
- PMID: 33103089
- PMCID: PMC7578749
- DOI: 10.1016/j.isci.2020.101654
Mitochondrial Protein Synthesis Is Essential for Terminal Differentiation of CD45- TER119-Erythroid and Lymphoid Progenitors
Abstract
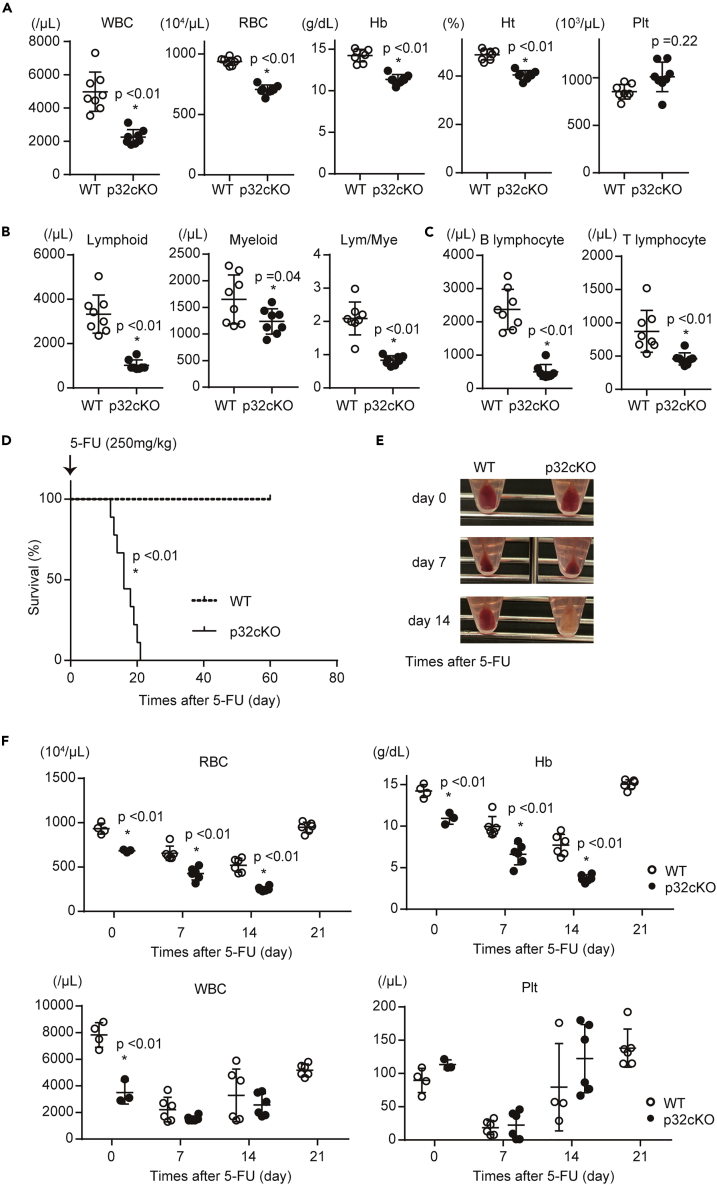
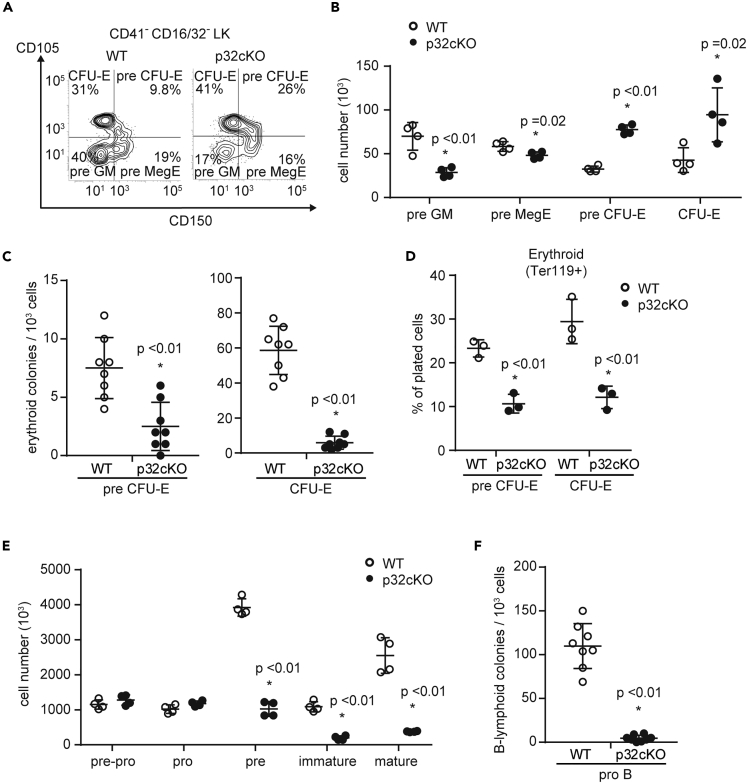
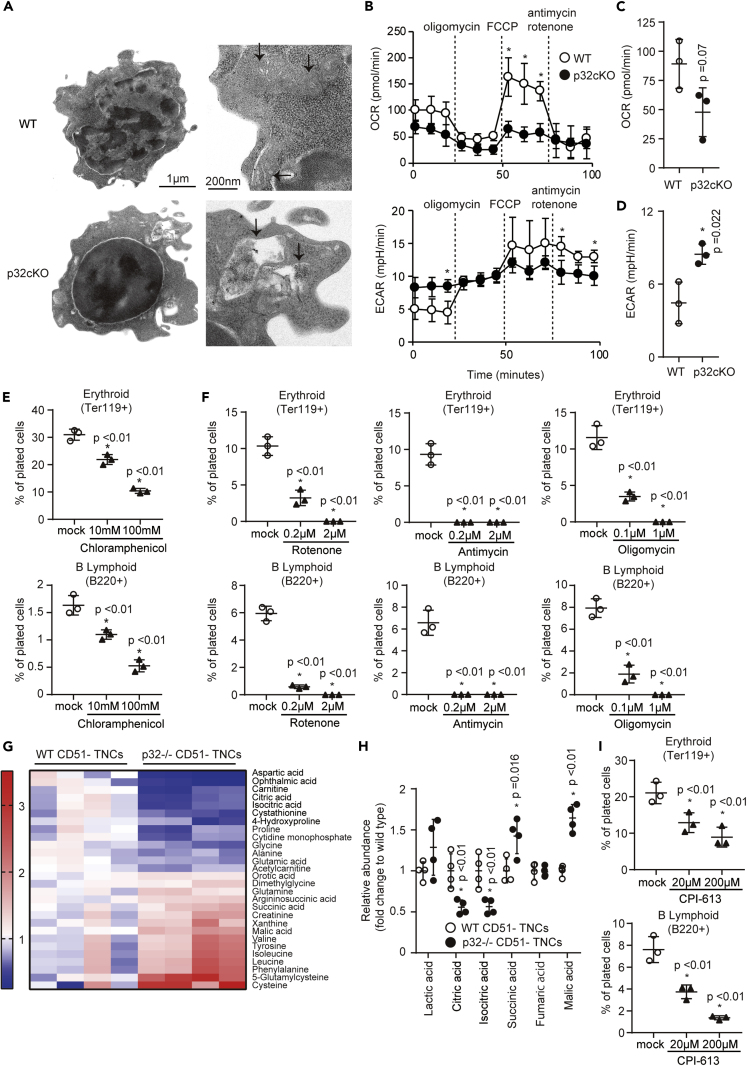
p32/C1qbp regulates mitochondrial protein synthesis and is essential for oxidative phosphorylation in mitochondria. Although dysfunction of p32/C1qbp impairs fetal development and immune responses, its role in hematopoietic differentiation remains unclear. Here, we found that mitochondrial dysfunction affected terminal differentiation of newly identified erythroid/B-lymphoid progenitors among CD45- Ter119- CD31- triple-negative cells (TNCs) in bone marrow. Hematopoietic cell-specific genetic deletion of p32/C1qbp (p32cKO) in mice caused anemia and B-lymphopenia without reduction of hematopoietic stem/progenitor cells. In addition, p32cKO mice were susceptible to hematopoietic stress with delayed recovery from anemia. p32/C1qbp-deficient CD51- TNCs exhibited impaired mitochondrial oxidation that consequently led to inactivation of mTORC1 signaling, which is essential for erythropoiesis. These findings uncover the importance of mitochondria, especially at the stage of TNCs during erythropoiesis, suggesting that dysregulation of mitochondrial protein synthesis is a cause of anemia and B-lymphopenia with an unknown pathology.
Keywords: Developmental Genetics; Molecular Biology.
© 2020 Kyushu University.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Cappelli E., Ravera S., Vaccaro D., Cuccarolo P., Bartolucci M., Panfoli I., Dufour C., Degan P. Mitochondrial respiratory complex I defects in Fanconi anemia. Trends Mol. Med. 2013;19:513–514. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
