

Macrophage ubiquitin-specific protease 2 contributes to motility, hyperactivation, capacitation, and in vitro fertilization activity of mouse sperm
- PMID: 33104844
- PMCID: PMC11073191
- DOI: 10.1007/s00018-020-03683-9
Macrophage ubiquitin-specific protease 2 contributes to motility, hyperactivation, capacitation, and in vitro fertilization activity of mouse sperm
Abstract
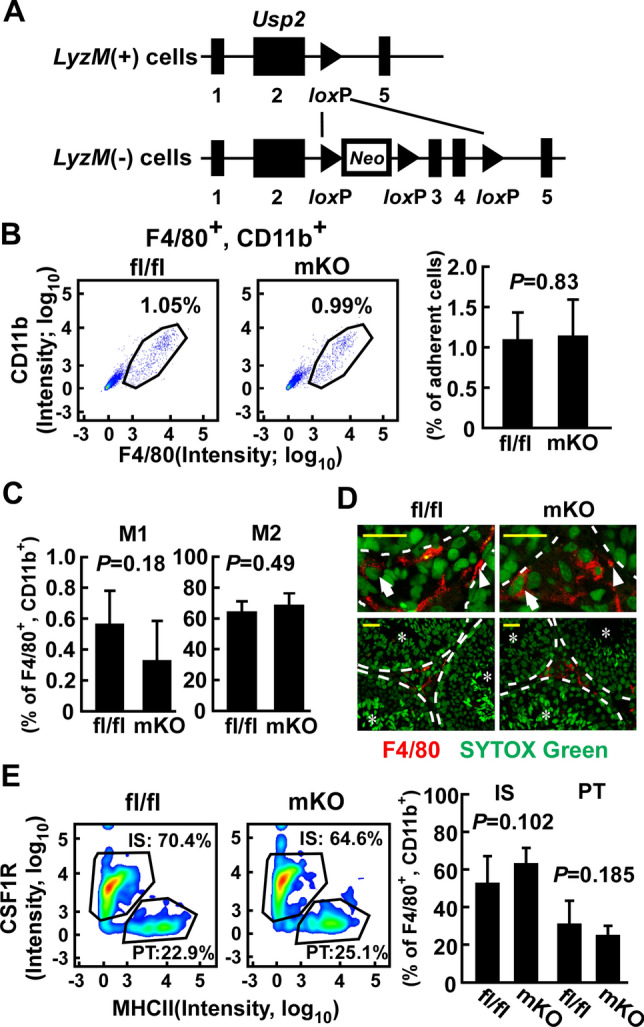
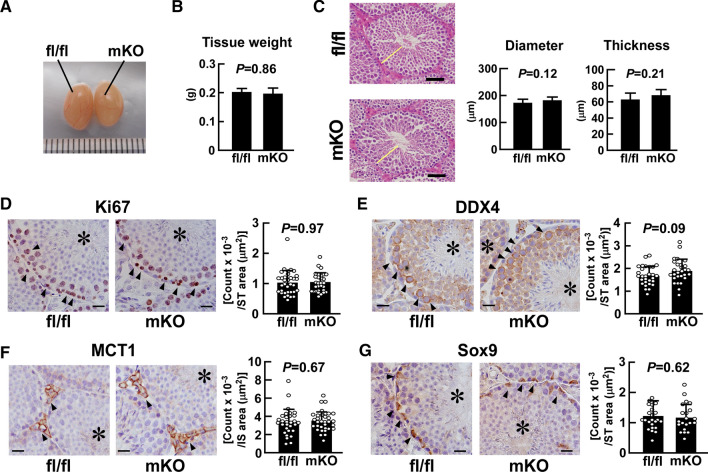
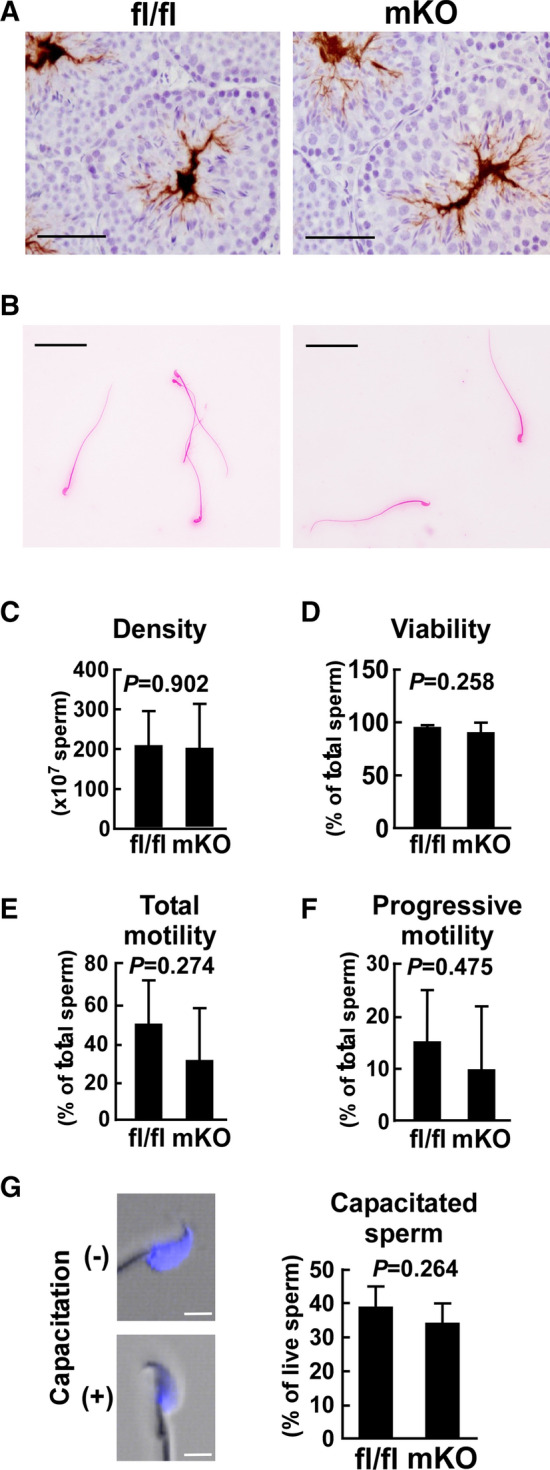
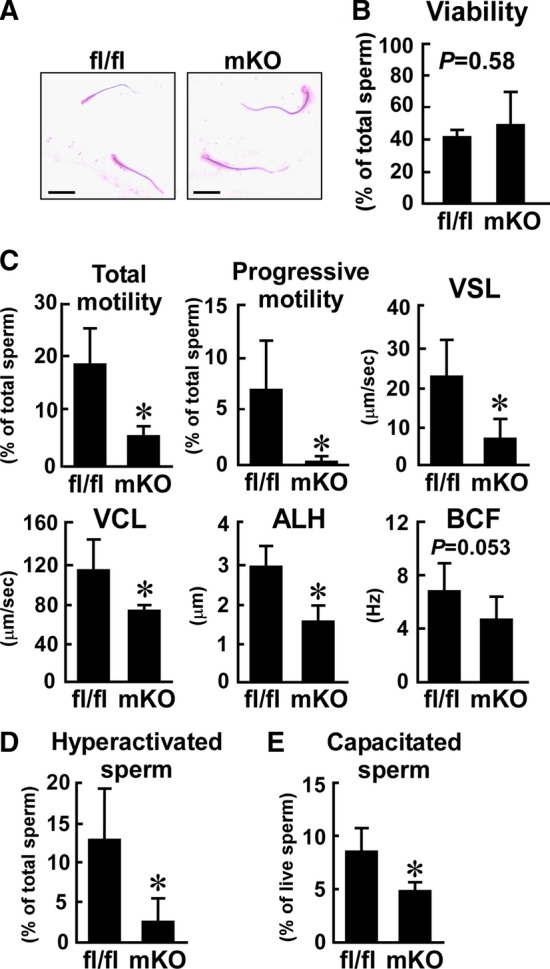
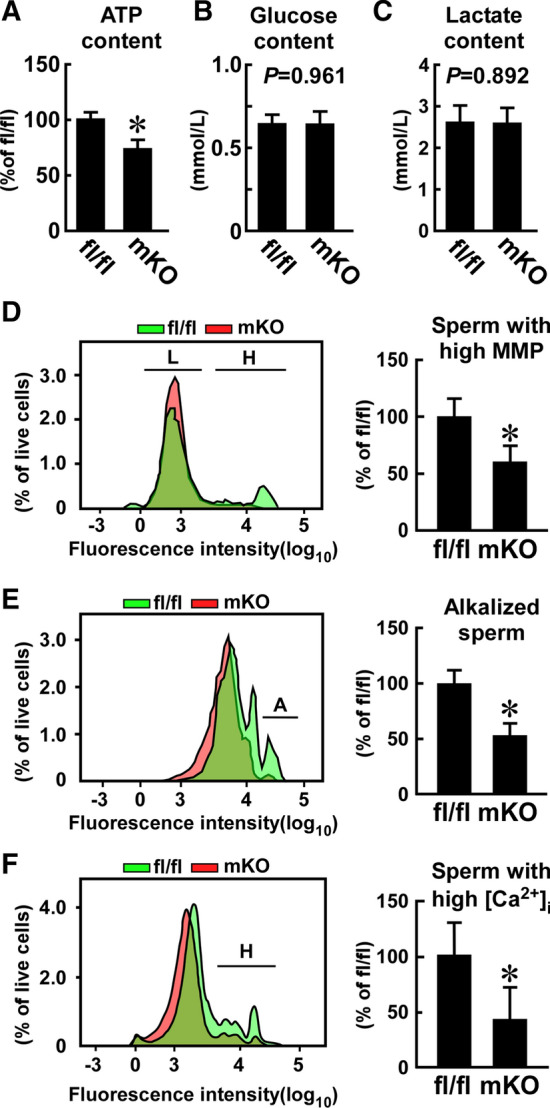
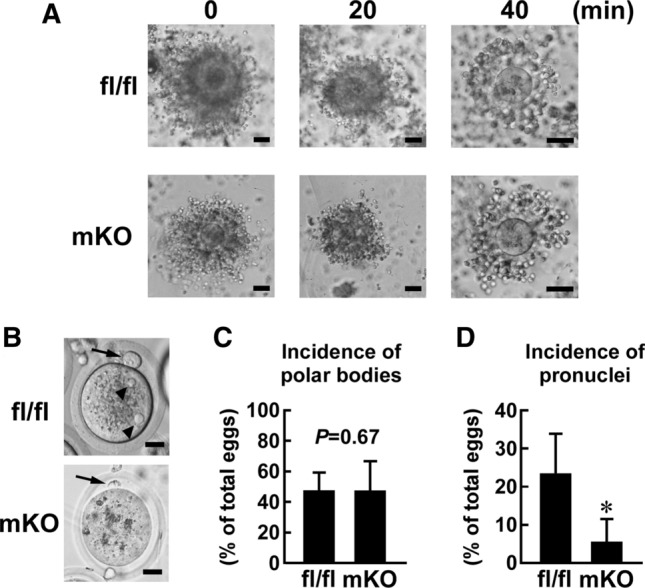
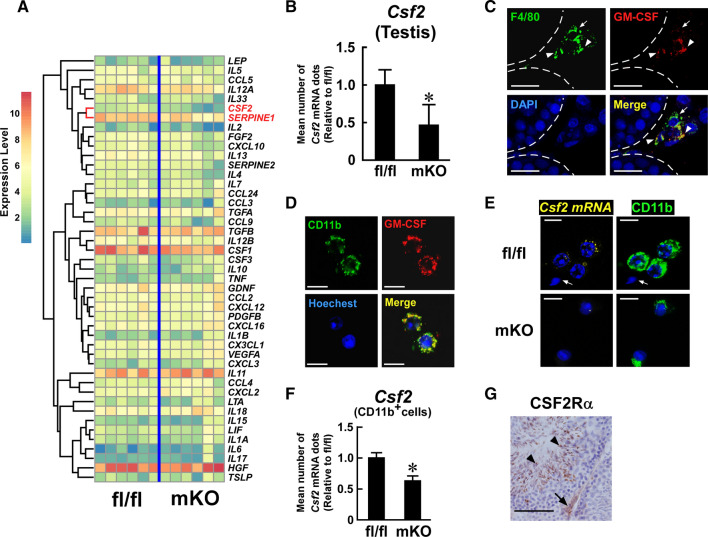
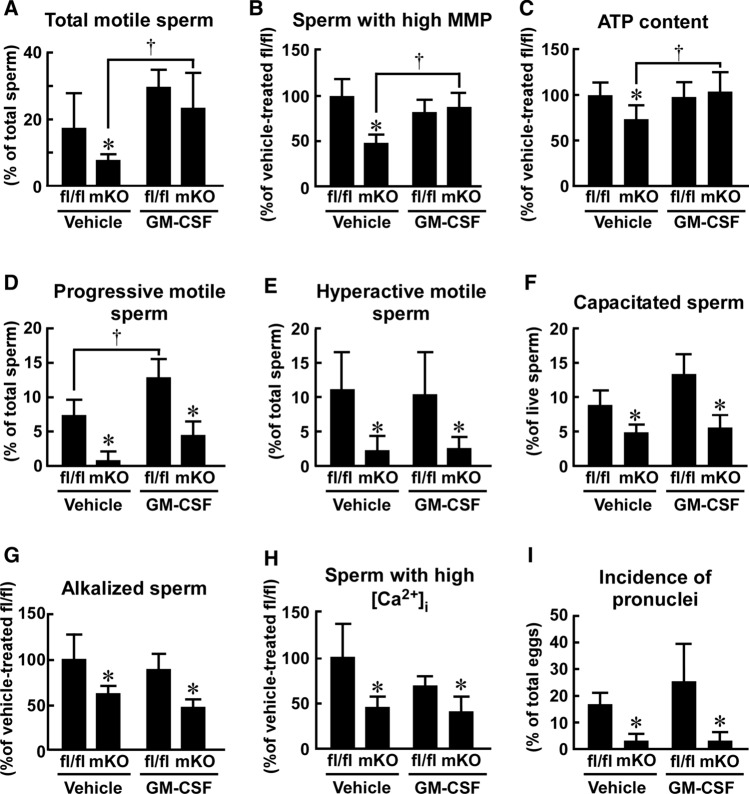
Macrophages are innate immune cells that contribute to classical immune functions and tissue homeostasis. Ubiquitin-specific protease 2 (USP2) controls cytokine production in macrophages, but its organ-specific roles are still unknown. In this study, we generated myeloid-selective Usp2 knockout (msUsp2KO) mice and specifically explored the roles of testicular macrophage-derived USP2 in reproduction. The msUsp2KO mice exhibited normal macrophage characteristics in various tissues. In the testis, macrophage Usp2 deficiency negligibly affected testicular macrophage subpopulations, spermatogenesis, and testicular organogenesis. However, frozen-thawed sperm derived from msUsp2KO mice exhibited reduced motility, capacitation, and hyperactivation. In addition, macrophage Usp2 ablation led to a decrease in the sperm population exhibiting high intracellular pH, calcium influx, and mitochondrial membrane potential. Interrupted pronuclei formation in eggs was observed when using frozen-thawed sperm from msUsp2KO mice for in vitro fertilization. Administration of granulocyte macrophage-colony stimulating factor (GM-CSF), whose expression was decreased in testicular macrophages derived from msUsp2KO mice, restored mitochondrial membrane potential and total sperm motility. Our observations demonstrate a distinct role of the deubiquitinating enzyme in organ-specific macrophages that directly affect sperm function.
Keywords: Capacitation; Granulocyte macrophage-colony stimulating factor; Male sterility; Myeloid cells; USP.
Conflict of interest statement
The authors have no conflicts of interest/competing interests to declare.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
