

A combinatorial native MS and LC-MS/MS approach reveals high intrinsic phosphorylation of human Tau but minimal levels of other key modifications
- PMID: 33106314
- PMCID: PMC7939451
- DOI: 10.1074/jbc.RA120.015882
A combinatorial native MS and LC-MS/MS approach reveals high intrinsic phosphorylation of human Tau but minimal levels of other key modifications
Abstract
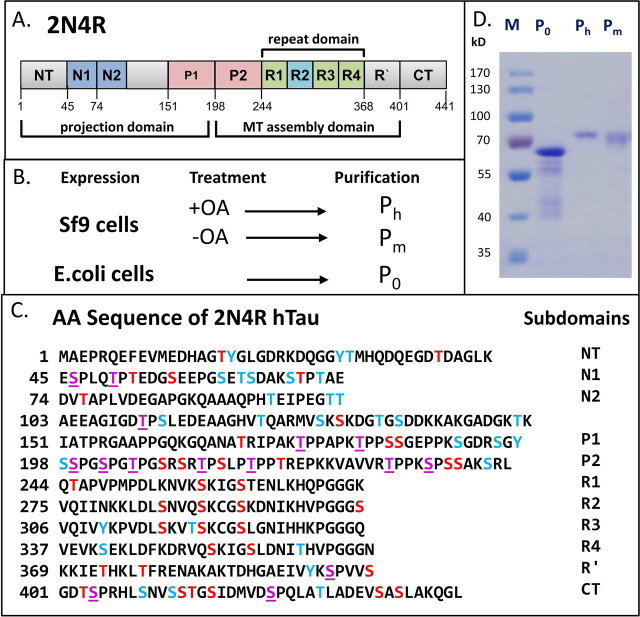
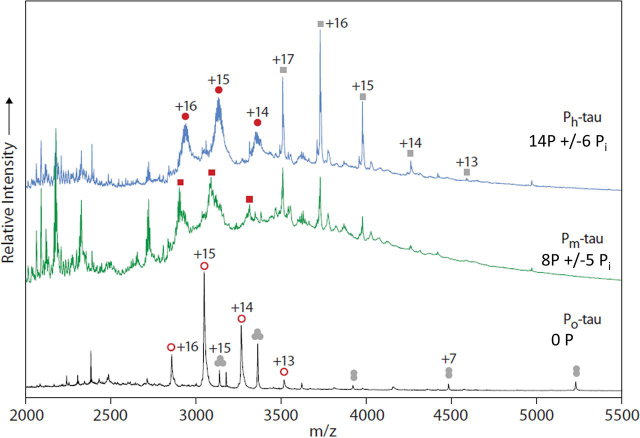
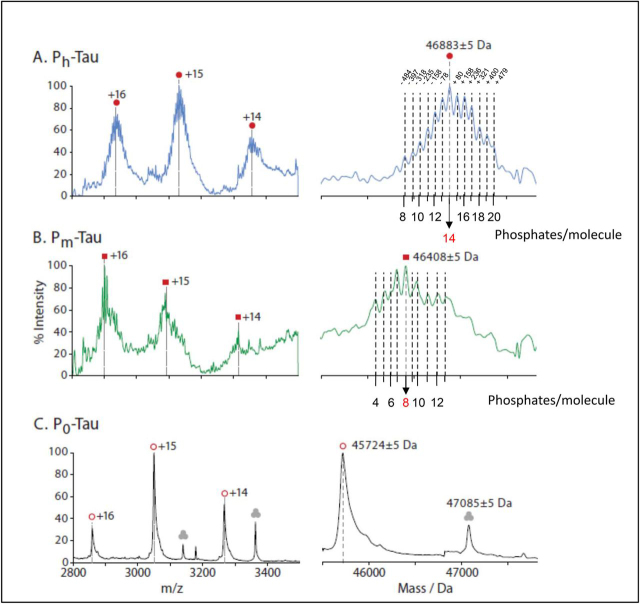
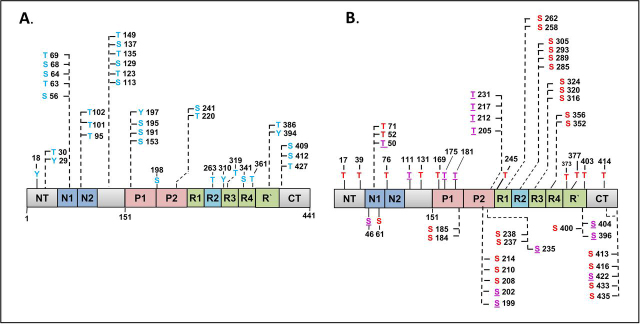
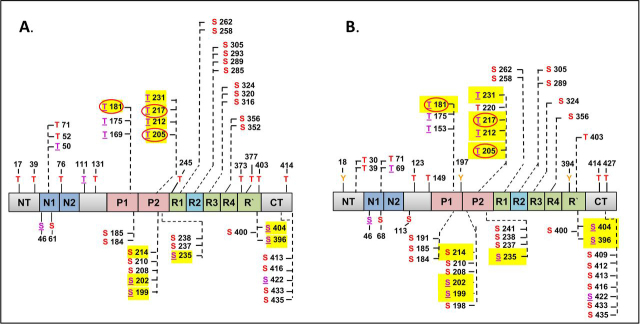
Abnormal changes of neuronal Tau protein, such as phosphorylation and aggregation, are considered hallmarks of cognitive deficits in Alzheimer's disease. Abnormal phosphorylation is thought to precede aggregation and therefore to promote aggregation, but the nature and extent of phosphorylation remain ill-defined. Tau contains ∼85 potential phosphorylation sites, which can be phosphorylated by various kinases because the unfolded structure of Tau makes them accessible. However, methodological limitations (e.g. in MS of phosphopeptides, or antibodies against phosphoepitopes) led to conflicting results regarding the extent of Tau phosphorylation in cells. Here we present results from a new approach based on native MS of intact Tau expressed in eukaryotic cells (Sf9). The extent of phosphorylation is heterogeneous, up to ∼20 phosphates per molecule distributed over 51 sites. The medium phosphorylated fraction Pm showed overall occupancies of ∼8 Pi (± 5) with a bell-shaped distribution; the highly phosphorylated fraction Ph had 14 Pi (± 6). The distribution of sites was highly asymmetric (with 71% of all P-sites in the C-terminal half of Tau). All sites were on Ser or Thr residues, but none were on Tyr. Other known posttranslational modifications were near or below our detection limit (e.g. acetylation, ubiquitination). These findings suggest that normal cellular Tau shows a remarkably high extent of phosphorylation, whereas other modifications are nearly absent. This implies that abnormal phosphorylations at certain sites may not affect the extent of phosphorylation significantly and do not represent hyperphosphorylation. By implication, the pathological aggregation of Tau is not likely a consequence of high phosphorylation.
Keywords: Alzheimer's disease; LC-MS; Tau protein (Tau); mass spectrometry (MS); native mass spectrometry; phosphorylation; protein aggregation.
© 2020 Drepper et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Grundke-Iqbal I., Iqbal K., Tung Y.C., Quinlan M., Wisniewski H.M., Binder L.I. Abnormal phosphorylation of the microtubule-associated protein tau (tau) in Alzheimer cytoskeletal pathology. Proc. Natl. Acad. Sci. U. S. A. 1986;83:4913–4917. doi: 10.1073/pnas.83.13.4913. 3088567. - DOI - PMC - PubMed
-
- DeVos S.L., Miller R.L., Schoch K.M., Holmes B.B., Kebodeaux C.S., Wegener A.J., Chen G., Shen T., Tran H., Nichols B., Zanardi T.A., Kordasiewicz H.B., Swayze E.E., Bennett C.F., Diamond M.I. Tau reduction prevents neuronal loss and reverses pathological tau deposition and seeding in mice with tauopathy. Sci. Transl. Med. 2017;9 doi: 10.1126/scitranslmed.aag0481. 28123067. - DOI - PMC - PubMed
-
- Yanamandra K., Kfoury N., Jiang H., Mahan T.E., Ma S., Maloney S.E., Wozniak D.F., Diamond M.I., Holtzman D.M. Anti-tau antibodies that block tau aggregate seeding in vitro markedly decrease pathology and improve cognition in vivo. Neuron. 2013;80:402–414. doi: 10.1016/j.neuron.2013.07.046. 24075978. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
