

ROS networks: designs, aging, Parkinson's disease and precision therapies
- PMID: 33106503
- PMCID: PMC7589522
- DOI: 10.1038/s41540-020-00150-w
ROS networks: designs, aging, Parkinson's disease and precision therapies
Abstract
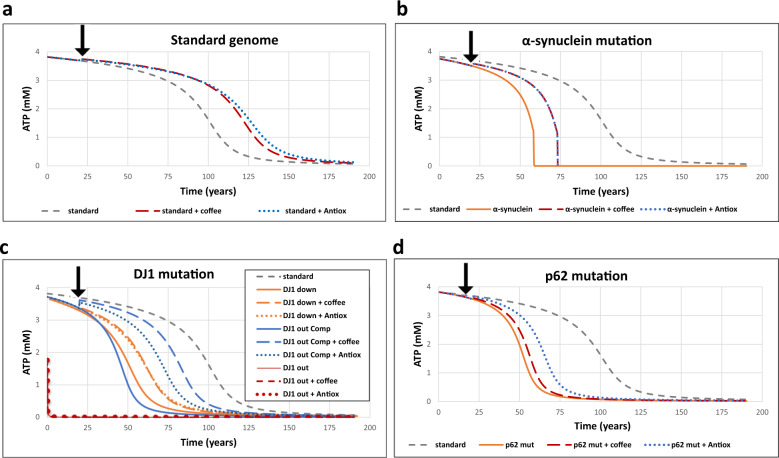
How the network around ROS protects against oxidative stress and Parkinson's disease (PD), and how processes at the minutes timescale cause disease and aging after decades, remains enigmatic. Challenging whether the ROS network is as complex as it seems, we built a fairly comprehensive version thereof which we disentangled into a hierarchy of only five simpler subnetworks each delivering one type of robustness. The comprehensive dynamic model described in vitro data sets from two independent laboratories. Notwithstanding its five-fold robustness, it exhibited a relatively sudden breakdown, after some 80 years of virtually steady performance: it predicted aging. PD-related conditions such as lack of DJ-1 protein or increased α-synuclein accelerated the collapse, while antioxidants or caffeine retarded it. Introducing a new concept (aging-time-control coefficient), we found that as many as 25 out of 57 molecular processes controlled aging. We identified new targets for "life-extending interventions": mitochondrial synthesis, KEAP1 degradation, and p62 metabolism.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Toxic Feedback Loop Involving Iron, Reactive Oxygen Species, α-Synuclein and Neuromelanin in Parkinson's Disease and Intervention with Turmeric.Mol Neurobiol. 2021 Nov;58(11):5920-5936. doi: 10.1007/s12035-021-02516-5. Epub 2021 Aug 23. Mol Neurobiol. 2021. PMID: 34426907 Review.
-
Dynamic modelling of protein and oxidative metabolisms simulates the pathogenesis of Parkinson's disease.IET Syst Biol. 2012 Jun;6(3):65-72. doi: 10.1049/iet-syb.2011.0075. IET Syst Biol. 2012. PMID: 22757585
-
Oxidative Modification and Its Implications for the Neurodegeneration of Parkinson's Disease.Mol Neurobiol. 2017 Mar;54(2):1404-1418. doi: 10.1007/s12035-016-9743-3. Epub 2016 Feb 3. Mol Neurobiol. 2017. PMID: 26843115 Review.
-
Oxidative stress involvement in alpha-synuclein oligomerization in Parkinson's disease cybrids.Antioxid Redox Signal. 2009 Mar;11(3):439-48. doi: 10.1089/ars.2008.2247. Antioxid Redox Signal. 2009. PMID: 18717628
-
The impact of reactive oxygen species and genetic mitochondrial mutations in Parkinson's disease.Gene. 2013 Dec 10;532(1):18-23. doi: 10.1016/j.gene.2013.07.085. Epub 2013 Aug 15. Gene. 2013. PMID: 23954870 Review.
Cited by
-
Novel insights into the mechanism of reactive oxygen species-mediated neurodegeneration.Neural Regen Res. 2023 Apr;18(4):746-749. doi: 10.4103/1673-5374.354509. Neural Regen Res. 2023. PMID: 36204830 Free PMC article. Review.
-
Dynamics of a model for the degradation mechanism of aggregated α-synuclein in Parkinson's disease.Front Comput Neurosci. 2023 Apr 13;17:1068150. doi: 10.3389/fncom.2023.1068150. eCollection 2023. Front Comput Neurosci. 2023. PMID: 37122994 Free PMC article.
-
Pasture intake protects against commercial diet-induced lipopolysaccharide production facilitated by gut microbiota through activating intestinal alkaline phosphatase enzyme in meat geese.Front Immunol. 2022 Dec 8;13:1041070. doi: 10.3389/fimmu.2022.1041070. eCollection 2022. Front Immunol. 2022. PMID: 36569878 Free PMC article.
-
Energy Metabolic Disorder of Astrocytes May Be an Inducer of Migraine Attack.Brain Sci. 2022 Jun 28;12(7):844. doi: 10.3390/brainsci12070844. Brain Sci. 2022. PMID: 35884650 Free PMC article.
-
Mitochondrial Phenotypes in Parkinson's Diseases-A Focus on Human iPSC-Derived Dopaminergic Neurons.Cells. 2021 Dec 7;10(12):3436. doi: 10.3390/cells10123436. Cells. 2021. PMID: 34943944 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/F003528/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/C008219/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G530225/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/I004696/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/I017186/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/I00470X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/I004688/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J500422/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J003883/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/J020060/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
