

Nfkb2 variants reveal a p100-degradation threshold that defines autoimmune susceptibility
- PMID: 33107914
- PMCID: PMC7595743
- DOI: 10.1084/jem.20200476
Nfkb2 variants reveal a p100-degradation threshold that defines autoimmune susceptibility
Abstract
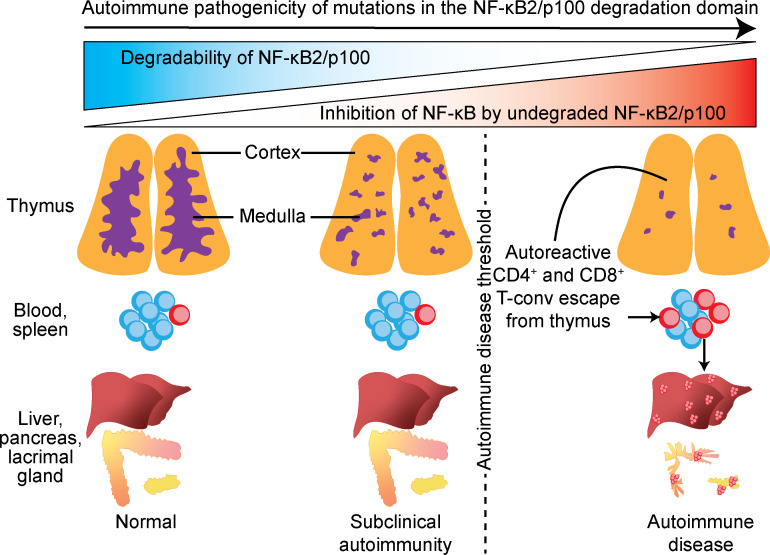
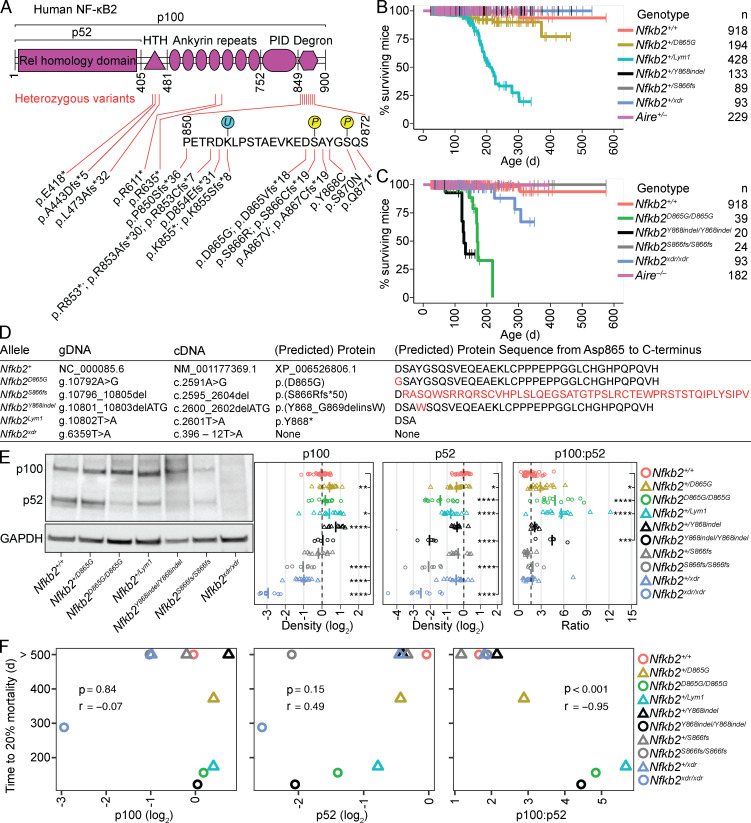
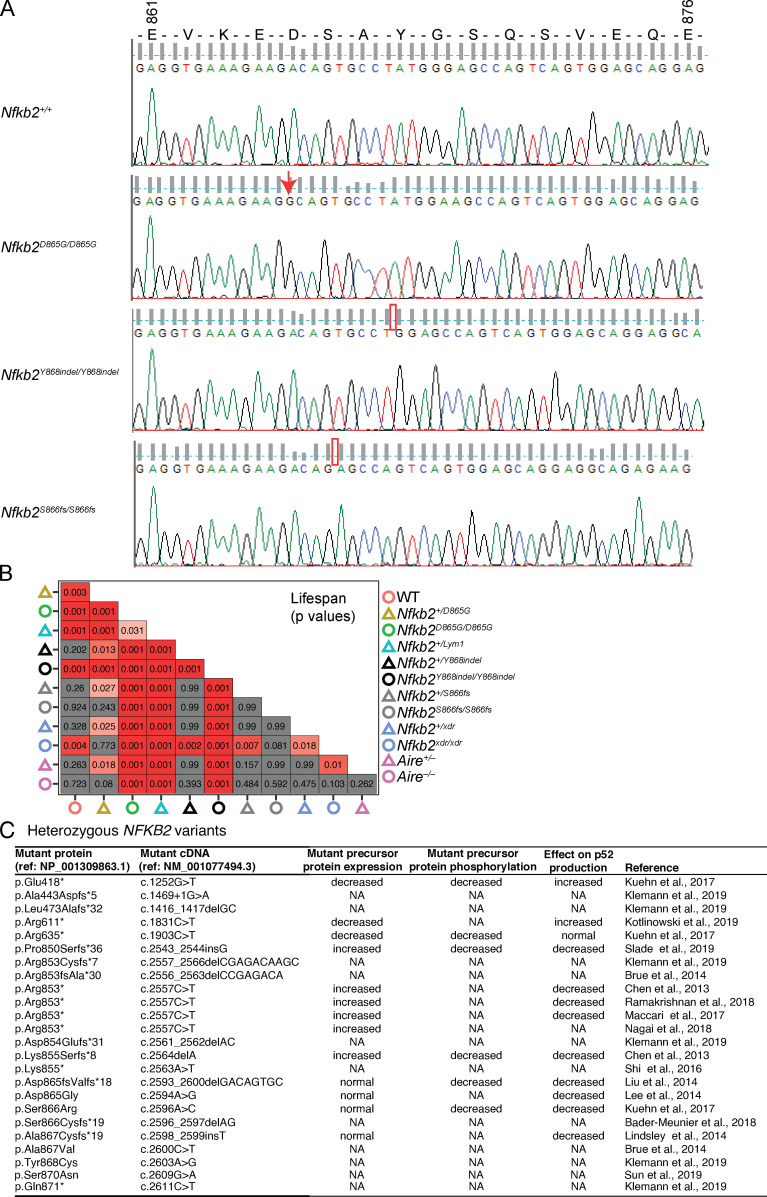
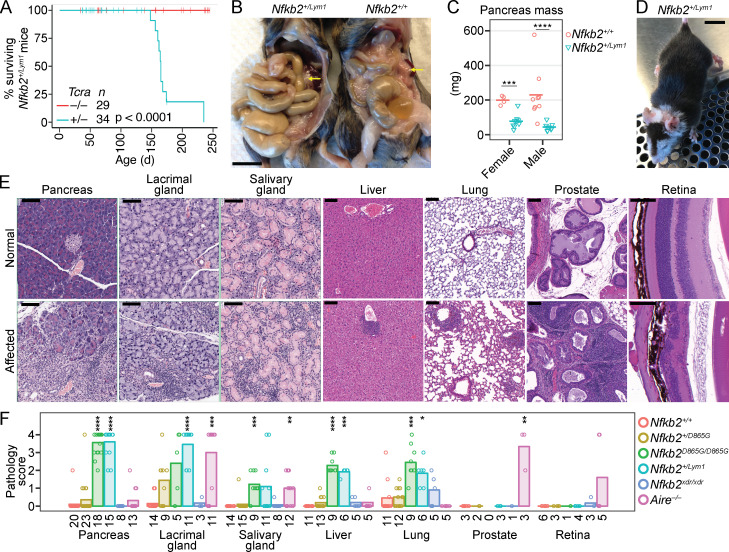
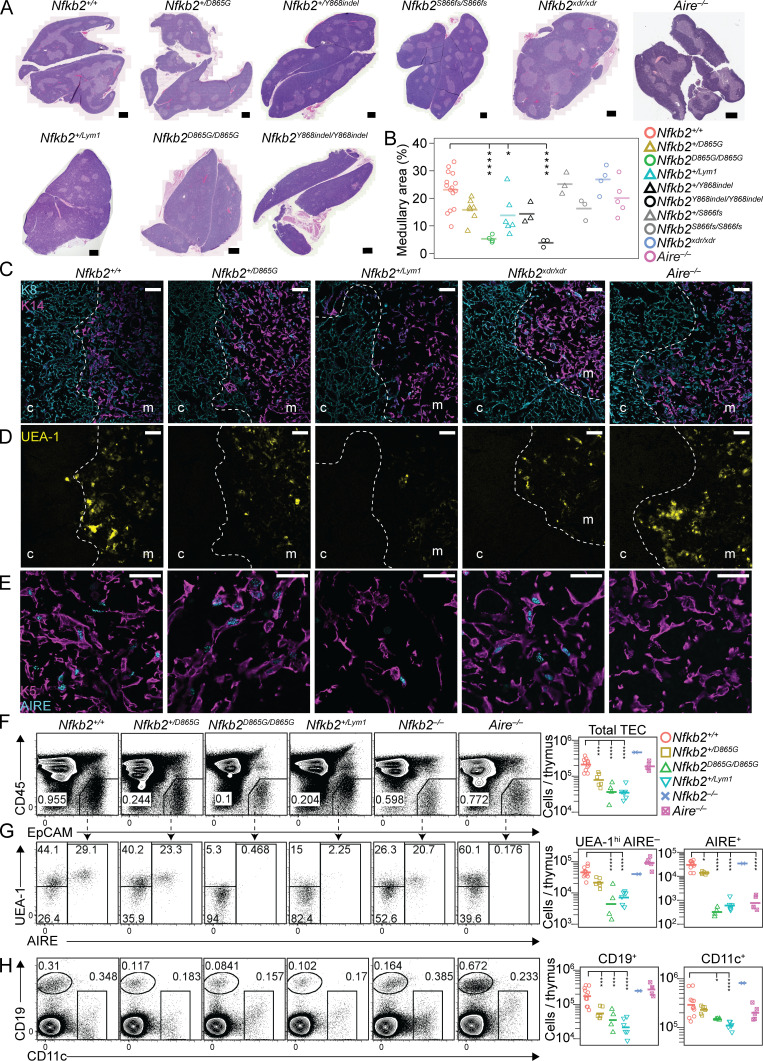
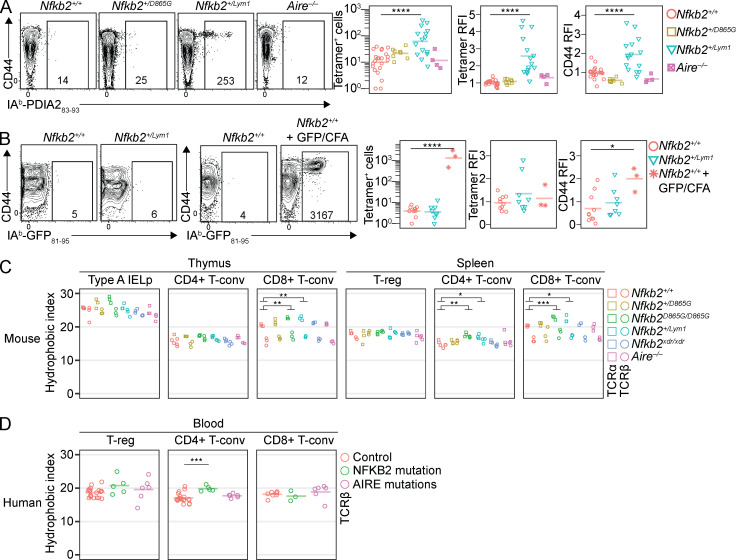
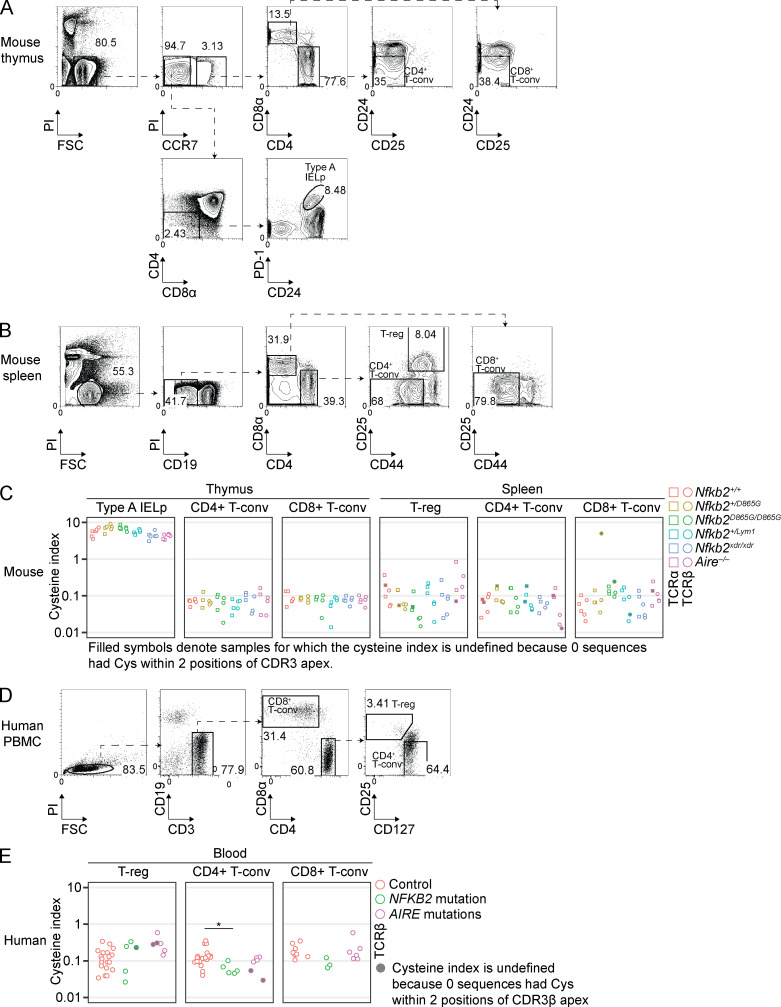
NF-κB2/p100 (p100) is an inhibitor of κB (IκB) protein that is partially degraded to produce the NF-κB2/p52 (p52) transcription factor. Heterozygous NFKB2 mutations cause a human syndrome of immunodeficiency and autoimmunity, but whether autoimmunity arises from insufficiency of p52 or IκB function of mutated p100 is unclear. Here, we studied mice bearing mutations in the p100 degron, a domain that harbors most of the clinically recognized mutations and is required for signal-dependent p100 degradation. Distinct mutations caused graded increases in p100-degradation resistance. Severe p100-degradation resistance, due to inheritance of one highly degradation-resistant allele or two subclinical alleles, caused thymic medullary hypoplasia and autoimmune disease, whereas the absence of p100 and p52 did not. We inferred a similar mechanism occurs in humans, as the T cell receptor repertoires of affected humans and mice contained a hydrophobic signature of increased self-reactivity. Autoimmunity in autosomal dominant NFKB2 syndrome arises largely from defects in nonhematopoietic cells caused by the IκB function of degradation-resistant p100.
© 2020 Wirasinha et al.
Conflict of interest statement
Disclosures: M.D. Keller reported "other" from Gilead outside the submitted work. D.H.D. Gray reported grants from Servier Pharmacetuicals outside the submitted work and is an employee of The Walter and Eliza Hall Institute of Medical Research, which receives milestone and royalty payments related to venetoclax (BCL-2 inhibitor). No other disclosures were reported.
Figures
Similar articles
-
Type-Specific Impacts of Protein Defects in Pathogenic NFKB2 Variants: Novel Clinical Findings From 138 Patients.J Allergy Clin Immunol Pract. 2025 Jan;13(1):192-201. doi: 10.1016/j.jaip.2024.10.015. Epub 2024 Oct 22. J Allergy Clin Immunol Pract. 2025. PMID: 39447838
-
Autoantibodies against type I IFNs in humans with alternative NF-κB pathway deficiency.Nature. 2023 Nov;623(7988):803-813. doi: 10.1038/s41586-023-06717-x. Epub 2023 Nov 8. Nature. 2023. PMID: 37938781 Free PMC article.
-
Gastric hyperplasia and increased proliferative responses of lymphocytes in mice lacking the COOH-terminal ankyrin domain of NF-kappaB2.J Exp Med. 1997 Oct 6;186(7):999-1014. doi: 10.1084/jem.186.7.999. J Exp Med. 1997. PMID: 9314550 Free PMC article.
-
How do nuclear factor kappa B (NF-κB)1 and NF-κB2 defects lead to the incidence of clinical and immunological manifestations of inborn errors of immunity?Expert Rev Clin Immunol. 2023 Mar;19(3):329-339. doi: 10.1080/1744666X.2023.2174105. Epub 2023 Feb 9. Expert Rev Clin Immunol. 2023. PMID: 36706462 Review.
-
Is NF-kappaB2/p100 a direct activator of programmed cell death?Cancer Cell. 2002 Dec;2(6):431-3. doi: 10.1016/s1535-6108(02)00213-1. Cancer Cell. 2002. PMID: 12498710 Review.
Cited by
-
Case report: novel NFKB2 variant associated with pediatric eosinophilic granulomatosis with polyangiitis (EGPA) in the COVID-19 pandemic.Pediatr Rheumatol Online J. 2025 Mar 31;23(1):35. doi: 10.1186/s12969-025-01086-3. Pediatr Rheumatol Online J. 2025. PMID: 40165201 Free PMC article.
-
Altered mRNA Expression of NFKB1 and NFKB2 Genes in Penile Lichen Sclerosus, Penile Cancer and Zoon Balanitis.J Clin Med. 2022 Dec 7;11(24):7254. doi: 10.3390/jcm11247254. J Clin Med. 2022. PMID: 36555871 Free PMC article.
-
Autoimmune Polyendocrinopathy-Candidiasis-Ectodermal Dystrophy.Front Pediatr. 2021 Nov 1;9:723532. doi: 10.3389/fped.2021.723532. eCollection 2021. Front Pediatr. 2021. PMID: 34790633 Free PMC article. Review.
-
Mistuned NF-κB signaling in lymphocytes: lessons from relevant inborn errors of immunity.Clin Exp Immunol. 2023 Apr 25;212(2):117-128. doi: 10.1093/cei/uxad006. Clin Exp Immunol. 2023. PMID: 36651621 Free PMC article.
-
[Development of autoimmunity in patients with primary immunodeficiencies].Rev Med Inst Mex Seguro Soc. 2023 Mar 1;61(2):189-195. Rev Med Inst Mex Seguro Soc. 2023. PMID: 37200522 Free PMC article. Spanish.
References
-
- Brue, T., Quentien M.H., Khetchoumian K., Bensa M., Capo-Chichi J.M., Delemer B., Balsalobre A., Nassif C., Papadimitriou D.T., Pagnier A., et al. . 2014. Mutations in NFKB2 and potential genetic heterogeneity in patients with DAVID syndrome, having variable endocrine and immune deficiencies. BMC Med. Genet. 15:139 10.1186/s12881-014-0139-9 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
