

Specificity in glycosylation of multiple flagellins by the modular and cell cycle regulated glycosyltransferase FlmG
- PMID: 33108275
- PMCID: PMC7591256
- DOI: 10.7554/eLife.60488
Specificity in glycosylation of multiple flagellins by the modular and cell cycle regulated glycosyltransferase FlmG
Abstract
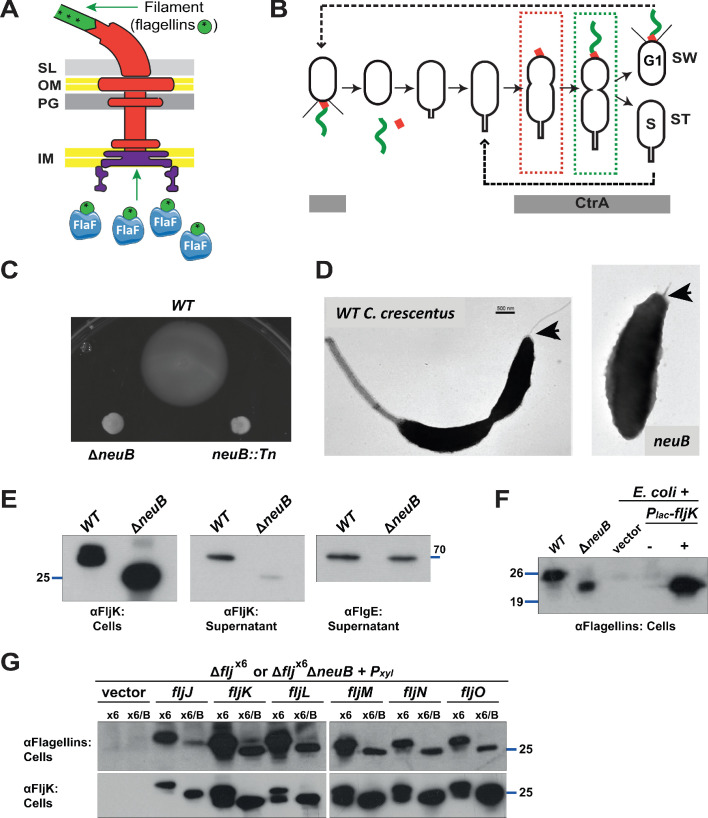
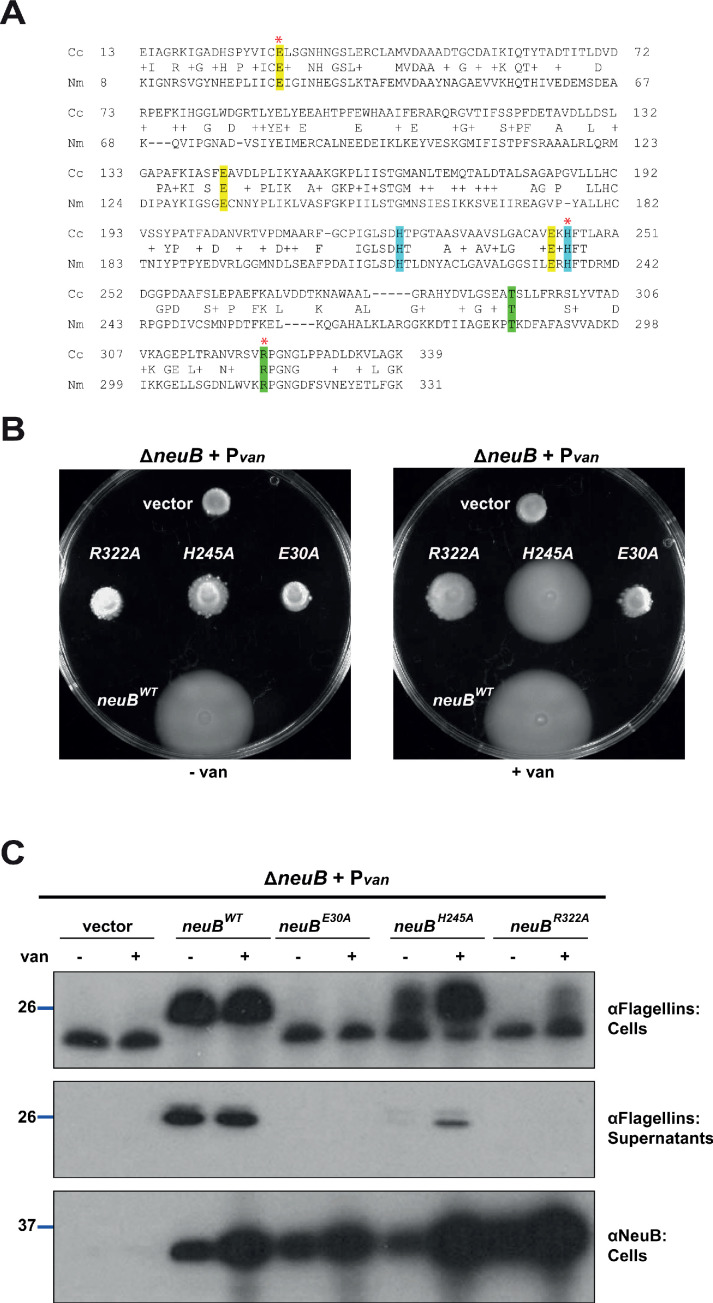
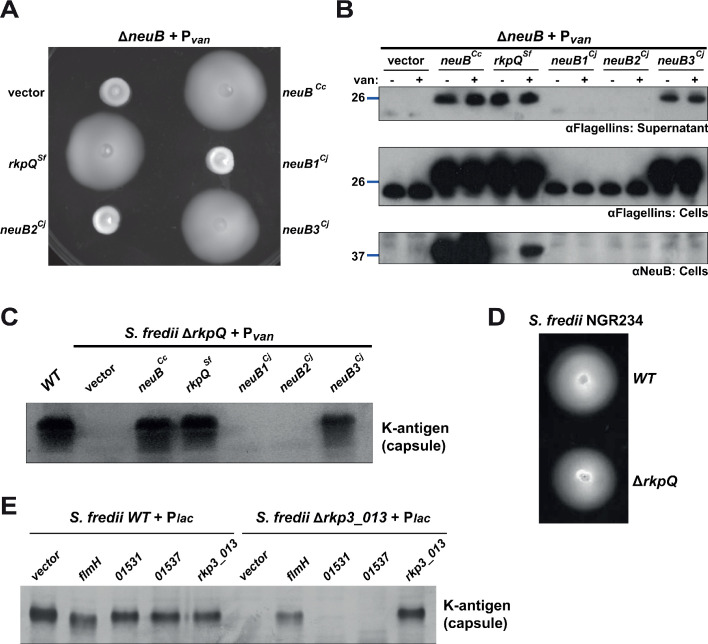
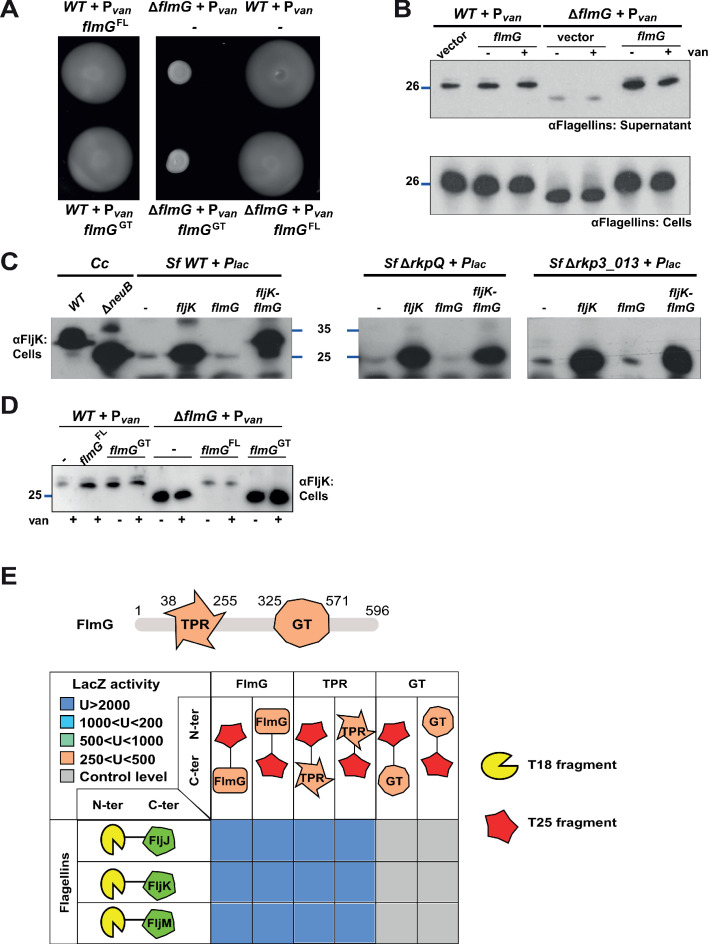
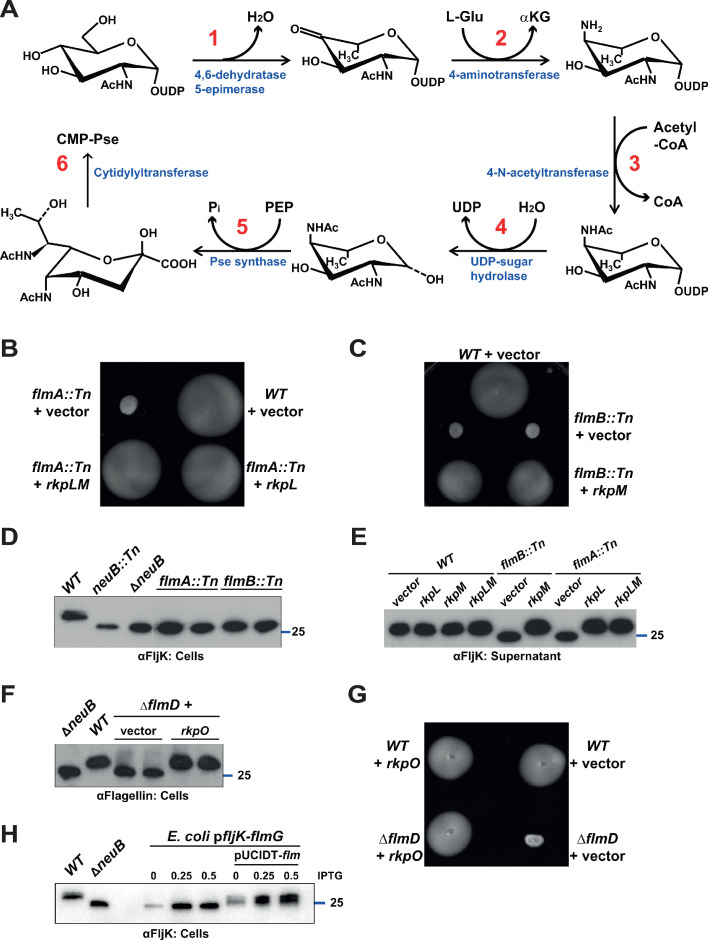
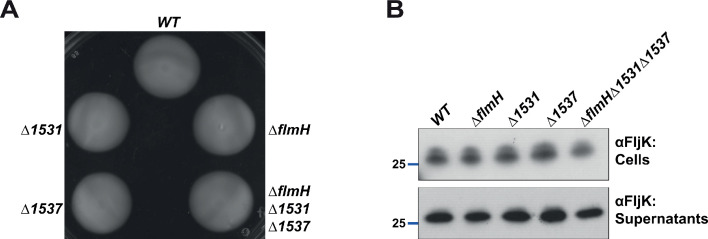
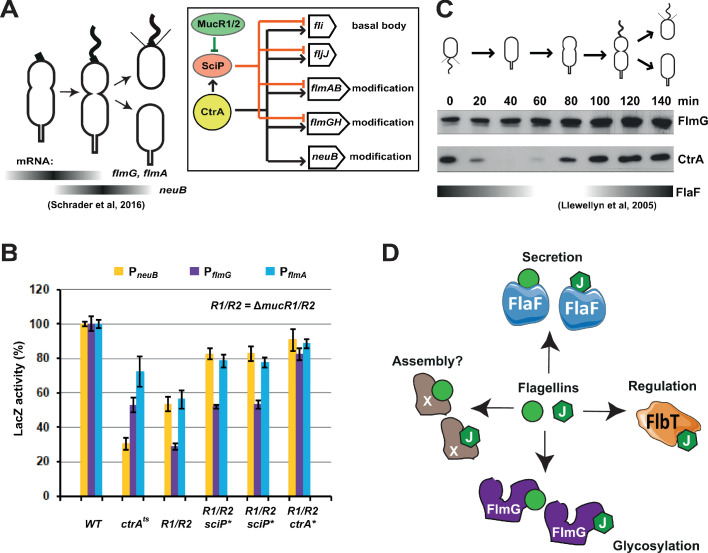
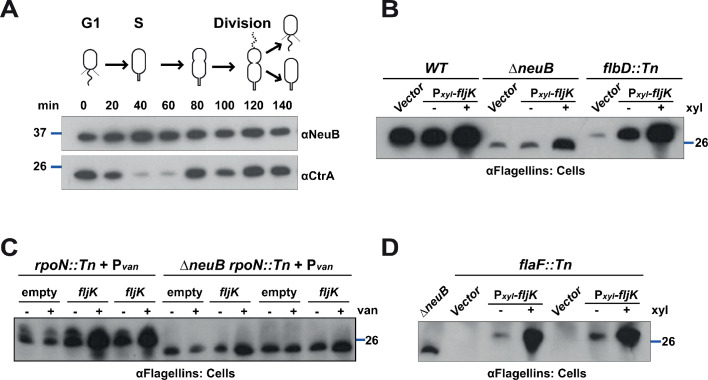
How specificity is programmed into post-translational modification of proteins by glycosylation is poorly understood, especially for O-linked glycosylation systems. Here we reconstitute and dissect the substrate specificity underpinning the cytoplasmic O-glycosylation pathway that modifies all six flagellins, five structural and one regulatory paralog, in Caulobacter crescentus, a monopolarly flagellated alpha-proteobacterium. We characterize the biosynthetic pathway for the sialic acid-like sugar pseudaminic acid and show its requirement for flagellation, flagellin modification and efficient export. The cognate NeuB enzyme that condenses phosphoenolpyruvate with a hexose into pseudaminic acid is functionally interchangeable with other pseudaminic acid synthases. The previously unknown and cell cycle-regulated FlmG protein, a defining member of a new class of cytoplasmic O-glycosyltransferases, is required and sufficient for flagellin modification. The substrate specificity of FlmG is conferred by its N-terminal flagellin-binding domain. FlmG accumulates before the FlaF secretion chaperone, potentially timing flagellin modification, export, and assembly during the cell division cycle.
Keywords: Caulobacter crescentus; FlmG; flagellin; flagellum; glycosylation; infectious disease; microbiology; pseudaminic acid.
© 2020, Ardissone et al.
Conflict of interest statement
SA, NK No competing interests declared, PV The authors declare a pending patent application PAT7460EP00 on FlmG-dependent soluble protein glycosylation systems in bacteria.
Figures
Similar articles
-
Stereoisomer-specific reprogramming of a bacterial flagellin sialyltransferase.EMBO J. 2023 Mar 1;42(5):e112880. doi: 10.15252/embj.2022112880. Epub 2023 Jan 13. EMBO J. 2023. PMID: 36636824 Free PMC article.
-
Specificity and modularity of flagellin nonulosonic acid glycosyltransferases.Trends Microbiol. 2022 Feb;30(2):109-111. doi: 10.1016/j.tim.2021.10.005. Epub 2021 Nov 13. Trends Microbiol. 2022. PMID: 34782242
-
Secretion Relieves Translational Co-repression by a Specialized Flagellin Paralog.Dev Cell. 2020 Nov 23;55(4):500-513.e4. doi: 10.1016/j.devcel.2020.10.005. Epub 2020 Oct 27. Dev Cell. 2020. PMID: 33113346
-
Flagellin glycosylation with pseudaminic acid in Campylobacter and Helicobacter: prospects for development of novel therapeutics.Cell Mol Life Sci. 2018 Apr;75(7):1163-1178. doi: 10.1007/s00018-017-2696-5. Epub 2017 Oct 27. Cell Mol Life Sci. 2018. PMID: 29080090 Free PMC article. Review.
-
Positional information during Caulobacter cell differentiation.Curr Opin Genet Dev. 1991 Oct;1(3):324-9. doi: 10.1016/s0959-437x(05)80295-3. Curr Opin Genet Dev. 1991. PMID: 1840888 Review.
Cited by
-
Whole genome characterization of thermophilic Campylobacter species isolated from dairy manure in small specialty crop farms of Northeast Ohio.Front Microbiol. 2023 Mar 21;14:1074548. doi: 10.3389/fmicb.2023.1074548. eCollection 2023. Front Microbiol. 2023. PMID: 37025625 Free PMC article.
-
Harnessing the acceptor substrate promiscuity of Clostridium botulinum Maf glycosyltransferase to glyco-engineer mini-flagellin protein chimeras.Commun Biol. 2024 Aug 21;7(1):1029. doi: 10.1038/s42003-024-06736-y. Commun Biol. 2024. PMID: 39169227 Free PMC article.
-
Flagella at the Host-Microbe Interface: Key Functions Intersect With Redundant Responses.Front Immunol. 2022 Mar 24;13:828758. doi: 10.3389/fimmu.2022.828758. eCollection 2022. Front Immunol. 2022. PMID: 35401545 Free PMC article. Review.
-
Flagellar Structures from the Bacterium Caulobacter crescentus and Implications for Phage ϕ CbK Predation of Multiflagellin Bacteria.J Bacteriol. 2021 Feb 8;203(5):e00399-20. doi: 10.1128/JB.00399-20. Print 2021 Feb 8. J Bacteriol. 2021. PMID: 33288623 Free PMC article.
-
Brucella and Its Hidden Flagellar System.Microorganisms. 2021 Dec 31;10(1):83. doi: 10.3390/microorganisms10010083. Microorganisms. 2021. PMID: 35056531 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
