

In crystallo screening for proline analog inhibitors of the proline cycle enzyme PYCR1
- PMID: 33109600
- PMCID: PMC7939384
- DOI: 10.1074/jbc.RA120.016106
In crystallo screening for proline analog inhibitors of the proline cycle enzyme PYCR1
Abstract
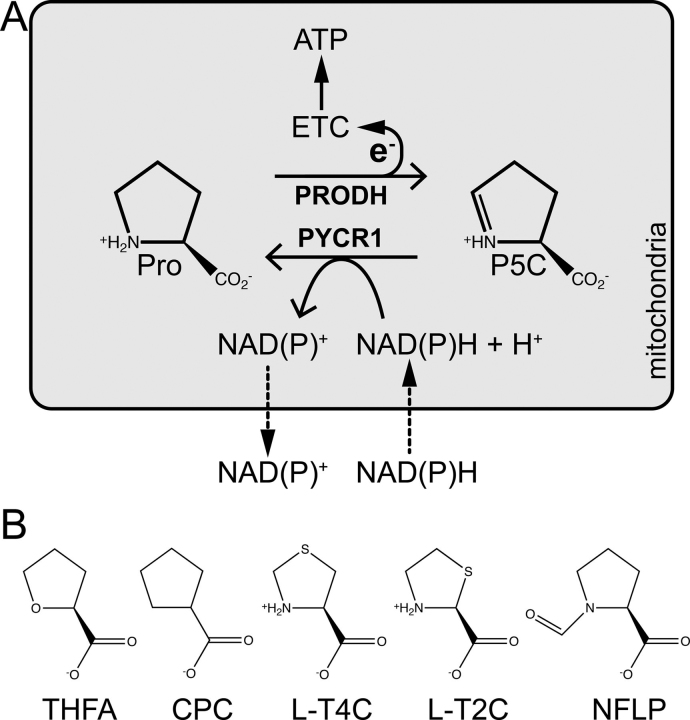
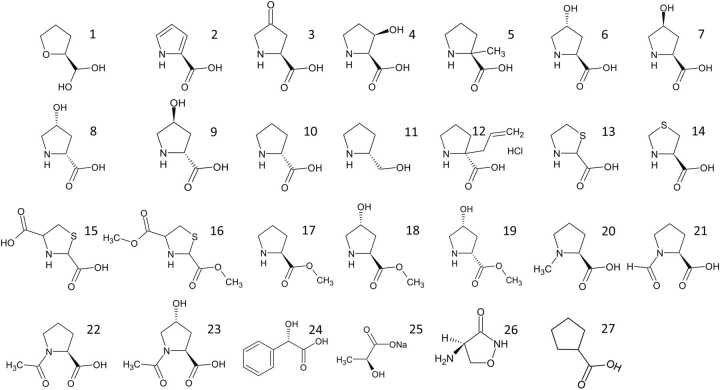
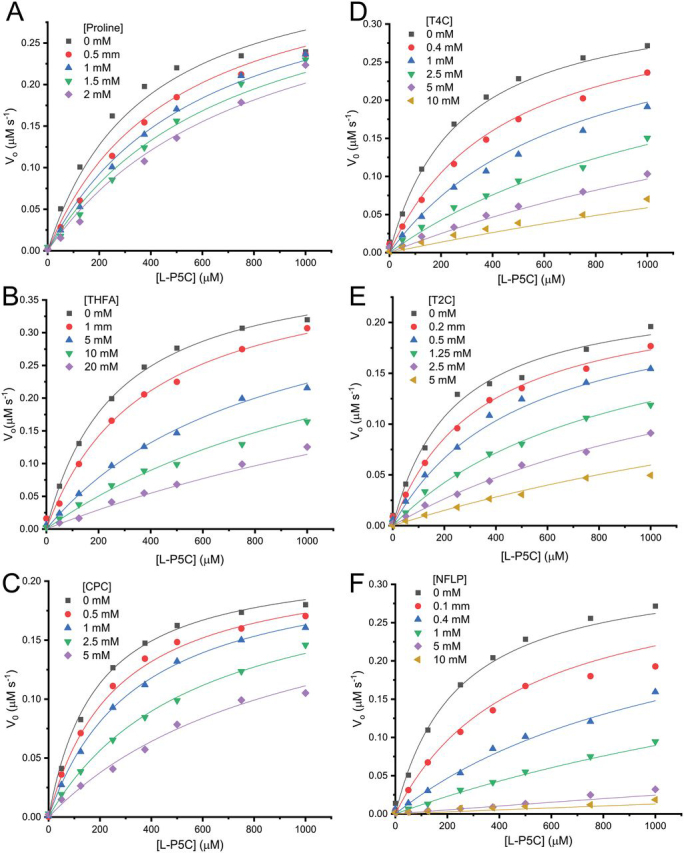
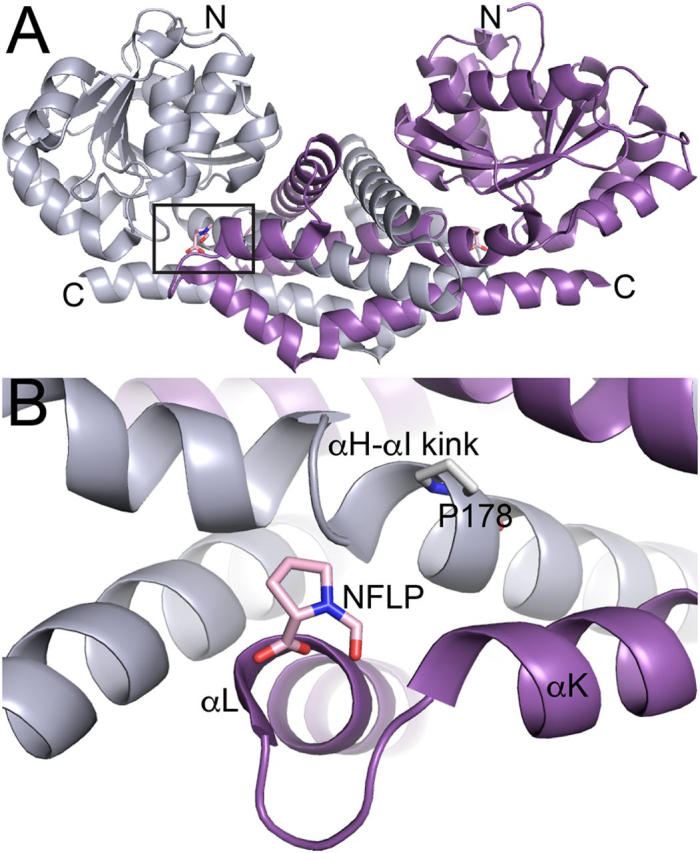
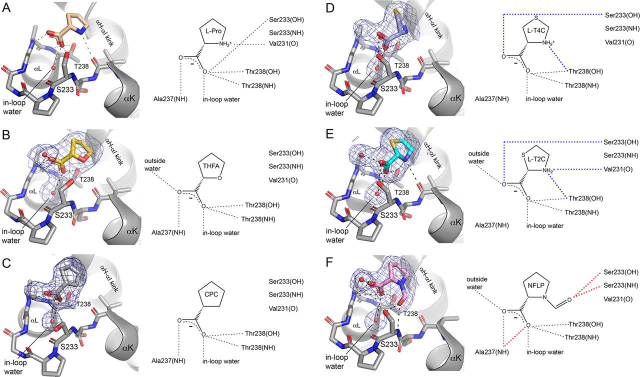
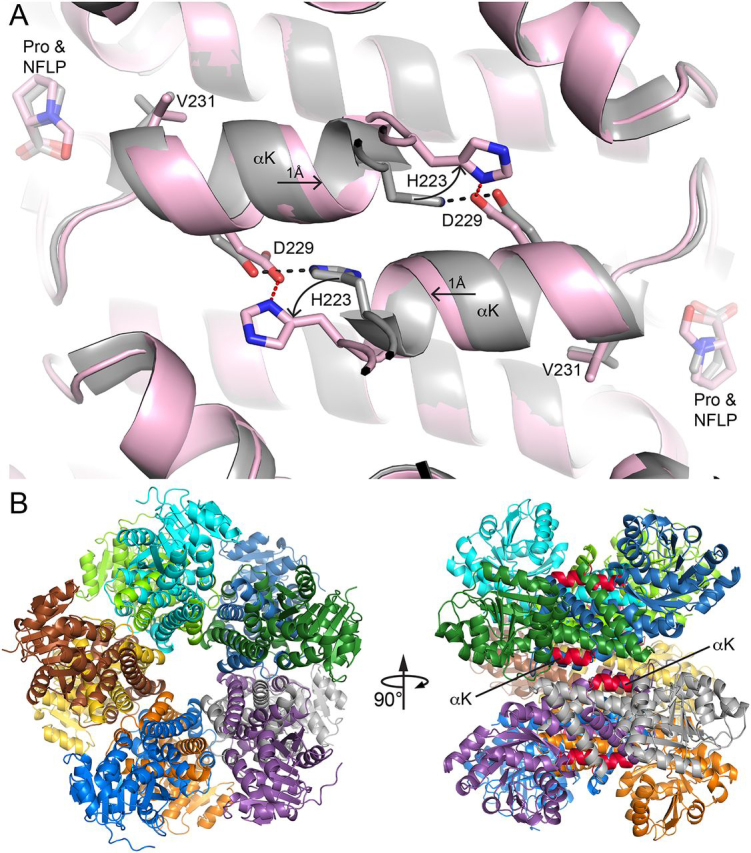
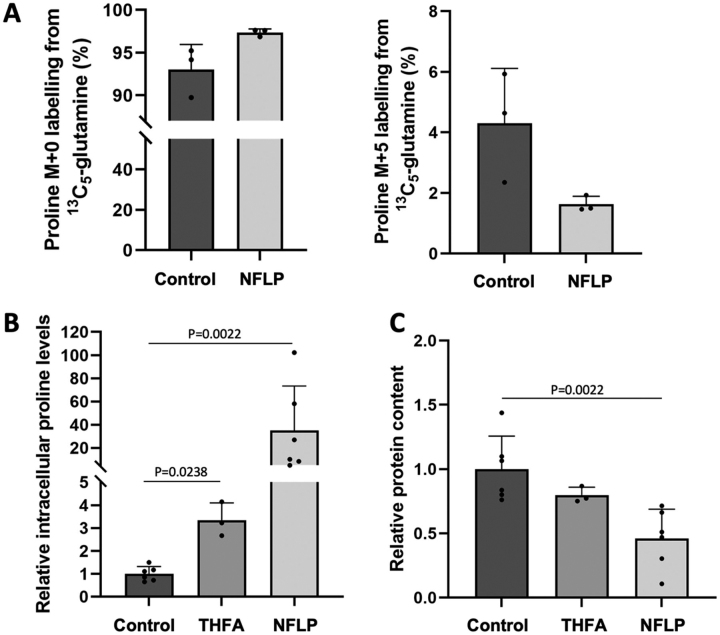
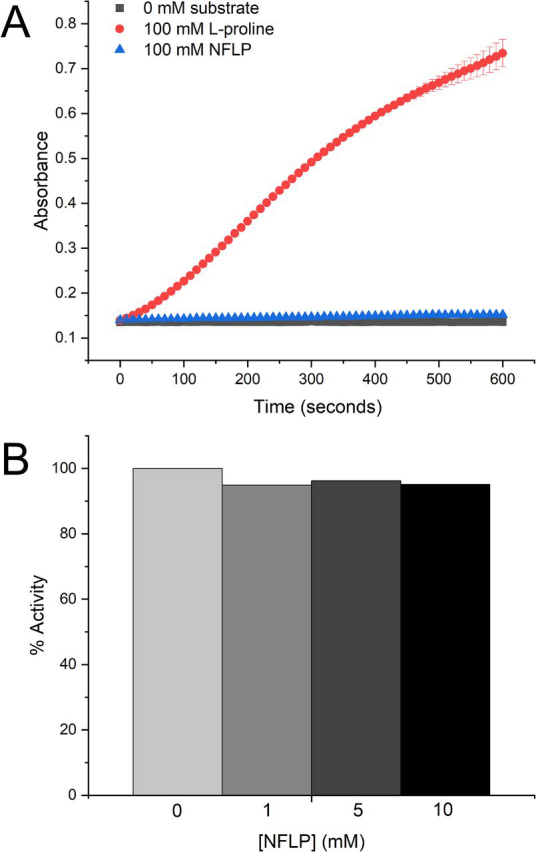
Pyrroline-5-carboxylate reductase 1 (PYCR1) catalyzes the biosynthetic half-reaction of the proline cycle by reducing Δ1-pyrroline-5-carboxylate (P5C) to proline through the oxidation of NAD(P)H. Many cancers alter their proline metabolism by up-regulating the proline cycle and proline biosynthesis, and knockdowns of PYCR1 lead to decreased cell proliferation. Thus, evidence is growing for PYCR1 as a potential cancer therapy target. Inhibitors of cancer targets are useful as chemical probes for studying cancer mechanisms and starting compounds for drug discovery; however, there is a notable lack of validated inhibitors for PYCR1. To fill this gap, we performed a small-scale focused screen of proline analogs using X-ray crystallography. Five inhibitors of human PYCR1 were discovered: l-tetrahydro-2-furoic acid, cyclopentanecarboxylate, l-thiazolidine-4-carboxylate, l-thiazolidine-2-carboxylate, and N-formyl l-proline (NFLP). The most potent inhibitor was NFLP, which had a competitive (with P5C) inhibition constant of 100 μm The structure of PYCR1 complexed with NFLP shows that inhibitor binding is accompanied by conformational changes in the active site, including the translation of an α-helix by 1 Å. These changes are unique to NFLP and enable additional hydrogen bonds with the enzyme. NFLP was also shown to phenocopy the PYCR1 knockdown in MCF10A H-RASV12 breast cancer cells by inhibiting de novo proline biosynthesis and impairing spheroidal growth. In summary, we generated the first validated chemical probe of PYCR1 and demonstrated proof-of-concept for screening proline analogs to discover inhibitors of the proline cycle.
Keywords: X-ray crystallography; breast cancer; enzyme inhibitor; enzyme kinetics; tumor metabolism.
© 2020 Christensen et al.
Conflict of interest statement
Conflict of interest—S.-M. F. has received funding from Bayer, Merck, and Black Belt Therapeutics and has consulted for Fund+.
Figures
Similar articles
-
Structure, biochemistry, and gene expression patterns of the proline biosynthetic enzyme pyrroline-5-carboxylate reductase (PYCR), an emerging cancer therapy target.Amino Acids. 2021 Dec;53(12):1817-1834. doi: 10.1007/s00726-021-02999-5. Epub 2021 May 18. Amino Acids. 2021. PMID: 34003320 Free PMC article. Review.
-
Screening a knowledge-based library of low molecular weight compounds against the proline biosynthetic enzyme 1-pyrroline-5-carboxylate 1 (PYCR1).Protein Sci. 2024 Jul;33(7):e5072. doi: 10.1002/pro.5072. Protein Sci. 2024. PMID: 39133178 Free PMC article.
-
Novel Fragment Inhibitors of PYCR1 from Docking-Guided X-ray Crystallography.J Chem Inf Model. 2024 Mar 11;64(5):1704-1718. doi: 10.1021/acs.jcim.3c01879. Epub 2024 Feb 27. J Chem Inf Model. 2024. PMID: 38411104 Free PMC article.
-
Resolving the cofactor-binding site in the proline biosynthetic enzyme human pyrroline-5-carboxylate reductase 1.J Biol Chem. 2017 Apr 28;292(17):7233-7243. doi: 10.1074/jbc.M117.780288. Epub 2017 Mar 3. J Biol Chem. 2017. PMID: 28258219 Free PMC article.
-
The key enzyme PYCR1 in proline metabolism: a dual driver of cancer progression and fibrotic remodeling.J Enzyme Inhib Med Chem. 2025 Dec;40(1):2545620. doi: 10.1080/14756366.2025.2545620. Epub 2025 Sep 2. J Enzyme Inhib Med Chem. 2025. PMID: 40891362 Free PMC article. Review.
Cited by
-
SENP3 affects the expression of PYCR1 to promote bladder cancer proliferation and EMT transformation by deSUMOylation of STAT3.Aging (Albany NY). 2022 Oct 11;14(19):8032-8045. doi: 10.18632/aging.204333. Epub 2022 Oct 11. Aging (Albany NY). 2022. PMID: 36227136 Free PMC article.
-
Structure, biochemistry, and gene expression patterns of the proline biosynthetic enzyme pyrroline-5-carboxylate reductase (PYCR), an emerging cancer therapy target.Amino Acids. 2021 Dec;53(12):1817-1834. doi: 10.1007/s00726-021-02999-5. Epub 2021 May 18. Amino Acids. 2021. PMID: 34003320 Free PMC article. Review.
-
Amino Acid Metabolic Vulnerabilities in Acute and Chronic Myeloid Leukemias.Front Oncol. 2021 Jul 1;11:694526. doi: 10.3389/fonc.2021.694526. eCollection 2021. Front Oncol. 2021. PMID: 34277440 Free PMC article. Review.
-
Expression and kinetic characterization of PYCR3.Arch Biochem Biophys. 2023 Jan 1;733:109468. doi: 10.1016/j.abb.2022.109468. Epub 2022 Nov 19. Arch Biochem Biophys. 2023. PMID: 36414121 Free PMC article.
-
Molecular and in vivo studies of a glutamate-class prolyl-endopeptidase for coeliac disease therapy.Nat Commun. 2022 Aug 1;13(1):4446. doi: 10.1038/s41467-022-32215-1. Nat Commun. 2022. PMID: 35915115 Free PMC article.
References
-
- Elia I., Broekaert D., Christen S., Boon R., Radaelli E., Orth M.F., Verfaillie C., Grünewald T.G.P., Fendt S.-M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017;8 doi: 10.1038/ncomms15267. 28492237. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
